Темой сегодняшнего разговора будет группа зеленящих стрептококков – наименование не очень легитимное, но прочно вошедшее в практику, как и обзывательство антибактериальной терапии – антибиотикотерапией, а кто мы такие, чтобы отменять традиции.
Под наименованием «Зеленящие стрептококки или S.viridans» прячется целая группа микроорганизмов, относящихся к альфа-зеленящим стрептококкам семейства Streptococcaceae. Эта группа является очень важным компонентом в составе нормально микрофлоры полости рта, слизистых оболочек дыхательных путей, пищеварительного тракта и половых органов. Поэтому обнаружив следующих ее представителей (S.gordonii, S.oralis, S. mutants, S.sanguis и S.salivarius) в посеве из полости рта, не надо немедленно начинать это лечить – этим вы только навредите организму, выбив из его состава важный компонент его нормальной микрофлоры.
На этом можно было бы и закончить, если бы не одно большое «но» — некоторые представители данного семейства умеют вызывать не только гнойные инфекции, но и являются причиной бактериального эндокардита не только у лиц с иммунодефицитами, но и у вполне здоровых пациентов, не следящих за состоянием собственных зубов. Поэтому если лаборатория присылает высеянный из крови зеленящий стрептококк у социально благополучного пациента, но с бактериальным эндокардитом, загляните ему в рот. Поверьте на слово, это не такая большая казуистика даже в наши дни, а антибактериальная терапия будет отличаться разительно от тех случаев, когда возбудителями выступают энтерококки или золотистый стафилококк.
Рассмотрим основных представителей семейства, на которых больше всего шансов наткнуться в антибиотикограммах.
Streptococcus sanguis – воистину могуч и вездесущ. Обнаруживается в зубных отложениях с самого прорезывания зубов и до конца жизни, где и есть основное место его обитания. Иногда может быть обнаружен на слизистой глотки и в фекалиях, куда попадают транзитом. При этом надо помнить, что этот микроб вносит существенный вклад в процессы, связанные с кариесом, занимая в последующем ведущее место в ассоциациях микроорганизмов, выделяемых из гнойного экссудата при воспалительных процессах в челюстно-лицевой области, включая одонтогенные абсцессы мозга и других органов. Кроме того, они же могут быть выделены их крови и сердечных клапанов больных инфекционным эндокардитом.
Streptococcus salivarius – также является одним из основных компонентов нормальной микрофлоры полости рта, а также, но в меньшем количестве, может быть выделен со слизистых дыхательного, желудочно-кишечного и урогенитального тракта. Особенно часто выделяются при анализе микрофлоры языка, слюны и фекалий. И крайне редко из крови при менингитах, септицимии и эндокардитах.
Streptococcus mutans – выделяется с поверхности зубов и фекалий. Этот микроб является кариесогенным, а значит, может в дальнейшем вызывать бактериемию и бактериальный эндокардит у пациентов, не занимающихся регулярной санацией кариозных очагов в полости рта.
Streptococcus mitis – в норме выделяется из слюны, мокроты фекалий, при этом может выделяться из верхних дыхательных путей и крови при нозокомиальном инфицировании.
Совсем неверно упоминать в этом блоке стрептококки группы Anginosus — Streptococcus intermedius/S.anginosus/S.constellatus, но так как мы обсуждаем в основном орофарингеальную флору, способную вызвать инфекционные эндокардиты (и чтобы не писать еще один, совсем малюсенький блок), мы упомянем их здесь. В норме они являются представителями пародонтогенной группы. Но стоит представителю группы S.intermedius оттуда выбраться, как он может вызывать абсцессы печени, мозга и эндокардиты, так как проявляет высокую тропность к их тканям.
Диагностические аспекты
Освоив вышеприведенный материал, мы видим, что только обнаружение данных микроорганизмов в стерильных средах может указывать на них как на потенциальных возбудителей. Почему все-таки потенциальных? Да потому, что в 80% случаев это может оказаться банальной контаминацией в связи с несоблюдением техники забора материала (все мы дышим, и если материал забирается с нарушением правил асептики и антисептики и без использования закрытых систем, то налететь в пробирку может много чего и не только из наших рта и носов). Таким образом, посевы нужно повторять.
Обнаружение этого же зоопарка в местах естественного его обитания не говорит нам практически ничего, если только пациент совсем не следит за полостью рта, и тогда ему можно настоятельно рекомендовать посещение стоматолога.
Аспекты антибактериальной терапии
Группа зеленящих стрептококков природно резистентна к тетрациклинам, макролидам и клиндомицину (до 50% штаммов), к биссептолу резистентно более 75%. Таким образом об этих группах мы даже не вспоминаем.
Само лечение должно быть комплексным и часто в сочетании с хирургическим. Выбор конкретных препаратов будет зависеть от локализации процесса, тяжести состояния пациента и выделенного возбудителя
Например, чувствительность
Streptococcus mitis будет выглядеть так (нажмите на картинку для увеличения):
А чувствительность к Streptococcus mutans – вот так (нажмите на картинку для увеличения):
Подробнее все рассмотреть можно по этой ссылке, ведь именно на этой базе данных работают автоматические и полуавтоматические анализаторы в микробиологических лабораториях.
Препаратами выбора будут являться:
Пенициллин в высоких дозировках (до 24 млн ЕД в сутки), возможно, в сочетании с аминогликозидами
Цефтриаксон
Ванкомицин
Как и в случае с любым стрептококком, крайне важно соблюдать длительность лечения, и она должна быть не менее 10-14 дней.
Исключение составляет бактериальный эндокардит, там сроки лечения как минимум удваиваются.
В заключение хотелось бы сказать, что как ни прискорбно это сознавать, но недоступность, в первую очередь финансовая, адекватной стоматологической помощи вновь привела к всплеску заболеваний связанных с неблагополучием в полости рта, таким образом, мы вновь вынуждены вспоминать, что инфекционный эндокардит – это не только болезнь наркоманов и лиц со сниженным иммунным статусом, но и тех, у кого недостаточно средств на хорошего стоматолога, а потому и нам не лишне иногда осматривать полость рта у пациентов с инфекциями кровотока или абсцессами в головном мозге или печени.
Поделиться с друзьями
Пожалуйста, заполните поля e-mail адресов и убедитесь в их правильности
Страница статьи : Российский педиатрический журнал
Doern CD, Burnham CA. It’s not easy being green: the viridans group streptococci, with a focus on pediatric clinical manifestations. J Clin Microbiol. 2010; 48(11): 3829-35. doi: 10.1128/JCM.01563-10.
Brenciani A, Tiberi E, Tili E, Mingoia M, Palmieri C, Varaldo PE, et al. Genetic determinants and elements associated with antibiotic resistance in viridans group streptococci. J Antimicrob Chemother. 2014; 69(5): 1197-204. doi: 10.1093/jac/dkt495.
Facklam R. What happened to the streptococci: overview of taxonomic and nomenclature changes. Clin Microbiol Rev. 2002; 15(4): 613-30.
Spellerberg B, Brandt C. Streptococcus. In: Versalovic J, Carroll KC, Funke G, Jorgensen JH, Landry ML and Warnock DW (ed.), Manual of Clinical Microbiology, 10th ed., ASM Press, Washington, DC, 2011; 331-49.
Póntigo F, Moraga M, Flores SV. Molecular phylogeny and a taxonomic proposal for the genus Streptococcus. Genet Mol Res. 2015; 14(3): 10905-18. doi: 10.4238/2015.
Маянский Н.А., Калакуцкая А.Н., Мотузова О.В., Ломинадзе Г.Г., Крыжановская О.А., Катосова Л.К. MALDI-TOF масс-спектрометрия в рутинной работе микробиологической лаборатории. Вопросы диагностики в педиатрии. 2011; 3(5): 20-5.
Angeletti S, Dicuonzo G, Avola A, Crea F, Dedej E, Vailati F. et al. Viridans Group Streptococci clinical isolates: MALDI-TOF mass spectrometry versus gene sequence-based identification. PLoS One. 2015; 10(3):e0120502. doi: 10.1371/journal.pone.0120502.
Harju I, Lange C, Kostrzewa M, Maier T, Rantakokko-Jalava K, Haanperä M. Improved Differentiation of Streptococcus pneumoniae and Other S. mitis Group Streptococci by MALDI Biotyper Using an Improved MALDI Biotyper Database Content and a Novel Result Interpretation Algorithm. J Clin Microbiol. 2017; 55(3): 914-22. doi: 10.1128/JCM.01990-16.
Su TY, Lee MH, Huang CT, Liu TP, Lu JJ. The clinical impact of patients with bloodstream infection with different groups of Viridans group streptococci by using matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS). Medicine (Baltimore). 2018; 97(50):e13607. doi: 10.1097/MD.0000000000013607.
Ioannidou S, Papaparaskevas J, Tassios PT, Foustoukou M, Legakis NJ, Vatopoulos AC. Prevalence and characterization of the mechanisms of macrolide, lincosamide and streptogramin resistance in viridans group streptococci. Int J Antimicrob Agents. 2003; 22(6): 626-9.
Ergin A, Ercis S, Hasçelik G. Macrolide resistance mechanisms and in vitro susceptibility patterns of viridans group streptococci isolated from blood cultures. J Antimicrob Chemother. 2006; 57(1): 139-41.
Chun S, Huh HJ, Lee NY. Species-specific difference in antimicrobial susceptibility among viridans group streptococci. Ann Lab Med. 2015; 35(2): 205-11. doi: 10.3343/alm.2015.35.2.205.
Маянский Н.А., Кварчия А.З., Пономаренко О.А., Лазарева А.В., Куличенко Т.В. Носительство оральных стрептококков, устойчивых к пенициллину и эритромицину, у детей с острыми респираторными инфекциями. Российский педиатрический журнал. 2018; 21(6): 337-44. DOI: http://dx.doi.org/10.18821/1560-9561-2018-21-6-337-44.
Mayanskiy N, Alyabieva N, Ponomarenko O, Pakhomov A, Kulichenko T, Ivanenko A. et al. Bacterial etiology of acute otitis media and characterization of pneumococcal serotypes and genotypes among children in Moscow, Russia. Pediatr Infect Dis J. 2015; 34(3): 255-60. doi: 10.1097/INF.0000000000000554.
Маянский Н. А., Алябьева Н.М., Пономаренко О.А., Куличенко Т.В., Артемова И.В., Лазарева А.В. и др. Динамика распространенности серотипов и антибиотикорезистентности носоглоточных пневмококков, выделенных у детей в 2010-2016 гг.: результаты ретроспективного когортного исследования. Вопросы современной педиатрии. 2017; 16 (5): 413-23. doi: 10.15690/vsp.v16i5.1806).
Rodriguez-Avial I, Rodriguez-Avial C, Culebras E, Picazo JJ. Distribution of tetracycline resistance genes tet(M), tet(O), tet(L) and tet(K) in blood isolates of viridans group streptococci harbouring erm(B) and mef(A) genes. Susceptibility to quinupristin/dalfopristin and linezolid. Int J Antimicrob Agents. 2003; 21(6): 536-41.
Smith A, Jackson MS, Kennedy H. Antimicrobial susceptibility of viridans group streptococcal blood isolates to eight antimicrobial agents. Scand J Infect Dis. 2004; 36(4): 259-63.
Rozkiewicz D, Daniluk T, Sciepuk M, Zaremba ML, Cylwik-Rokicka D, Luczaj-Cepowicz E et al. Prevalence rate and antibiotic susceptibility of oral viridans group streptococci (VGS) in healthy children population. Adv Med Sci. 2006; 51 Suppl 1: 191-5.
Nielsen MJ, Claxton S, Pizer B, Lane S, Cooke RP, Paulus S, Carrol ED. Viridans Group Streptococcal Infections in Children After Chemotherapy or Stem Cell Transplantation: A 10-year Review From a Tertiary Pediatric Hospital. Medicine (Baltimore). 2016; 95(9): e2952. doi: 10.1097/MD.0000000000002952.
Süzük S, Kaşkatepe B, Çetin M. Antimicrobial susceptibility against penicillin, ampicillin and vancomycin of viridans group Streptococcus in oral microbiota of patients at risk of infective endocarditis. Infez Med. 2016; 24(3): 190-3.
Hakenbeck R, Brückner R, Denapaite D, Maurer P. Molecular mechanisms of β-lactam resistance in Streptococcus pneumoniae. Future Microbiol. 2012; 7: 395-410.
Chi F, Nolte O, Bergmann C, Ip M, Hakenbeck R. Crossing the barrier: evolution and spread of a major class of mosaic pbp2x in S. pneumoniae, S. mitis and S oralis. Int J Med Microbiol. 2007; 297: 503-12. https://doi.org/10.1016/j.ijmm.
Jensen A, Valdorsson O, Frimodt-Moller N, Hollingshead S, Kilian M. Commensal streptococci serve as a reservoir for beta-lactam resistance genes in Streptococcus pneumoniae. Antimicrob Agents Chemother.2015; 59: 3529 -40. https://doi.org/10.1128/AAC.00429-15.
Leclercq R. Mechanisms of resistance to macrolides and lincosamides: nature of the resistance elements and their clinical implications. Clin Infect Dis. 2002; 34: 482-92.
Schroeder MR, Stephens DS. Macrolide Resistance in Streptococcus pneumoniae. Front Cell Infect Microbiol. 2016; 6: 98.
van der Linden M, Otten J, Bergmann C, Latorre C, Liñares J, Hakenbeck R. Insight into the Diversity of Penicillin-Binding Protein 2x Alleles and Mutations in Viridans Streptococci. Antimicrob Agents Chemother. 2017; 61(5). pii: e02646-16. doi: 10.1128/AAC.02646-16.
Катосова Л.К., Лазарева А.В., Хохлова Т.А., Пономаренко О.А., Алябьева Н.М. Распространение и механизмы устойчивости к макролидам Streptococcus pyogenes, выделенных у детей. Антибиотики и химиотерапия. 2016; 61(3-4): 23-9.
Shaikh N, Leonard E, Martin JM. Prevalence of streptococcal pharyngitis and streptococcal carriage in children: a meta-analysis. Pediatrics. 2010; 126(3): e557-64. doi: 10.1542/peds.2009-2648.
Streptococcus mitis qroup s s 10*6 — Вопрос лору
Если вы не нашли нужной информации среди ответов на этот вопрос, или же ваша проблема немного отличается от представленной, попробуйте задать дополнительный вопрос врачу на этой же странице, если он будет по теме основного вопроса. Вы также можете задать новый вопрос, и через некоторое время наши врачи на него ответят. Это бесплатно. Также можете поискать нужную информацию в похожих вопросах на этой странице или через страницу поиска по сайту. Мы будем очень благодарны, если Вы порекомендуете нас своим друзьям в социальных сетях.
Медпортал 03online.com осуществляет медконсультации в режиме переписки с врачами на сайте. Здесь вы получаете ответы от реальных практикующих специалистов в своей области. В настоящий момент на сайте можно получить консультацию по 71 направлению: специалиста COVID-19, аллерголога, анестезиолога-реаниматолога, венеролога, гастроэнтеролога, гематолога, генетика, гепатолога, гериатра, гинеколога, гинеколога-эндокринолога, гомеопата, дерматолога, детского гастроэнтеролога, детского гинеколога, детского дерматолога, детского инфекциониста, детского кардиолога, детского лора, детского невролога, детского нефролога, детского офтальмолога, детского психолога, детского пульмонолога, детского ревматолога, детского уролога, детского хирурга, детского эндокринолога, дефектолога, диетолога, иммунолога, инфекциониста, кардиолога, клинического психолога, косметолога, логопеда, лора, маммолога, медицинского юриста, нарколога, невропатолога, нейрохирурга, неонатолога, нефролога, нутрициолога, онколога, онкоуролога, ортопеда-травматолога, офтальмолога, паразитолога, педиатра, пластического хирурга, проктолога, психиатра, психолога, пульмонолога, ревматолога, рентгенолога, репродуктолога, сексолога-андролога, стоматолога, трихолога, уролога, фармацевта, физиотерапевта, фитотерапевта, флеболога, фтизиатра, хирурга, эндокринолога.
Мы отвечаем на 97.47% вопросов.
Оставайтесь с нами и будьте здоровы!
Посев на бета-гемолитический стрептококк группы В (Streptococcus group В, S. agalactiae)
Метод определения
Исследование проводят методом посева на плотные питательные среды. Идентификацию микроорганизмов проводят методом масс-спектрометрии с помощью прибора Microflex Brucker Daltonik MALDI Biotyper, BRUKER, Германия.
Исследуемый материал
Мазок из входа во влагалище или аноректальной области
Доступен выезд на дом
Синонимы: Streptococcus group В (S. agalactiae) Culture.
Краткое описание исследования «Посев на бета-гемолитический стрептококк группы В (Streptococcus group В, S.
agalactiae)»
Стрептококк группы В (СГВ, GBS, Streptococcus agalactiae) вызывает тяжело протекающие заболевания у новорожденных детей и представляет опасность для определенных пациентов других возрастных групп.
В подавляющем большинстве случаев новорожденные инфицируются СГВ во время родов от матери, причем чаще это происходит при вагинальном родоразрешении. СГВ вызывает ранние неонатальные инфекции, такие как сепсис, менингит, пневмония, остеомиелит, артрит и пиелонефрит, частота которых в разных странах колеблется от 0,2 до 5 и более на 1000 живорожденных детей. В акушерской практике со стрептококком группы В связывают бактериемию, инфекции мочевых путей, хориоамнионит, преждевременное излитие околоплодных вод, преждевременные роды, послеродовой эндометрит и др.
СГВ в организме человека могут колонизировать ротоглотку, анальную область прямой кишки, влагалище (чаще преддверие), урогенитальный тракт, кожу.
Установлено, что СГВ у 5-35% женщин можно обнаружить во влагалище и часто в уретре их половых партнеров Показана достоверная связь носительства СГВ во влагалище не только с ранними септицемиями новорожденных, но и с самопроизвольными выкидышами, преждевременными родами, преждевременным излитием околоплодных вод, мочевой инфекцией у беременных, рождением детей с низкой массой тела, развитием хориоамнионита в родах, эндометритом и сепсисом у родильниц. Наиболее высок уровень колонизации у женщин репродуктивного возраста и беременных женщин. У беременных СГВ выделяют из перечисленных областей в 7-30% случаев (чаще у молодых, имеющих высокую половую активность или пользующихся внутриматочными средствами контрацепции). Из них 60-75% остаются СГВ-носителями до конца беременности.
Для выявления колонизации влагалища беременных женщин стрептококком группы В проводится сбор вагинально-ректальных мазков. Во всех остальных случаях биоматериал берется только при наличии клинических признаков инфекции.
СГВ обладает природной чувствительностью к бета-лактамным антибиотикам, включая препараты пенициллинового ряда. Определение чувствительности необходимо в тех случаях, когда у пациентки имеет место индивидуальная непереносимость препаратов данной группы.
Выделяемые микроорганизмы и возбудители:
Streptococcus agalactiae.
С какой целью проводят посев на бета-гемолитический стрептококк группы В (Streptococcus group В, S.
agalactiae)
Посев на бета-гемолитический стрептококк группы В (S. agalactiae) предназначен для обследования беременных на 35-37-й неделе беременности с целью выявления бактерионосительства и определения необходимости проведения антибиотикопрофилактики в родах, для предупреждения неонатальных осложнений (септицемии, пневмонии и менингита новорожденных).
Что может повлиять на результат теста «Посев на бета-гемолитический стрептококк группы В (Streptococcus group В, S. agalactiae)»
Несоблюдение правил подготовки к исследованию может повлиять на результат теста.
Платформа геномных ресурсов и анализа группы устных Streptococcus mitis
Функции базы данных и встроенные инструменты биоинформатики
The S . mitis Виды группы являются важными колонизаторами ротовой полости и иногда связаны с серьезными инфекциями [15]. Кроме того, недавно было высказано предположение, что эти организмы играют важную роль в патогенезе гриппа [8]. Таким образом, геномное исследование разнообразных S . mitis Бактерии группы необходимы для понимания того, как эти микроорганизмы переходят от комменсального образа жизни во рту к последующему патогенезу.Однако для широкого спектра S не существует специализированной базы данных генома. mitis групповых геномов для сравнительной геномики. Хотя большинство баз данных по биологическим геномам сосредоточены только на содержании генома и генетической изменчивости, мы определили необходимость создания функциональных биоинформатических инструментов для исследования детерминант вирулентности в геномах посредством сравнительной патогеномики, а также для сравнения содержания генома и генетической изменчивости в пределах S . mitis группа бактерий.
Инструмент парного сравнения генома (PGC)
Мы разработали и настроили веб-инструмент PGC для S . mitis группирует бактерии, что позволяет пользователям выбирать и проводить попарные сравнения между двумя выбранными пользователем геномами Streptococcus . Список геномов Streptococcus доступен в инструменте PGC StreptoBase, что позволяет пользователям выбрать два генома Streptococcus для сравнения штаммов или видов.Кроме того, пользователи могут загрузить свои собственные последовательности генома, нуклеотидов или белков, и сравнить их с геномами Streptococcus в StreptoBase.
Вкратце, конвейер PGC поддерживается NUCmer, который разработан для выравнивания полногеномных последовательностей, и Circos, который является хорошо зарекомендовавшим себя инструментом для визуализации генома. После того, как пользователи отправят свои задания на наш сервер, PGC вызовет программу NUCmer для согласования выбранных пользователем геномов, а внутренние сценарии будут использоваться для обработки выходных данных выравнивания генома и создания входных файлов, анализируемых в Circos, чтобы сгенерировать макет круговой идеограммы выравнивания. В отличие от обычного линейного отображения выравниваний, круговая компоновка показывает взаимосвязь между парами позиций с кариотипами и связями, кодирующими положение, размер и ориентацию связанных геномных элементов.
В веб-интерфейсе PGC предусмотрены три определяемых пользователем параметра, включая минимальный процент идентичности (%), порог слияния (bp) и порог связи (bp). Граница минимального процента идентичности определяет гомологичную область (представленную связями / лентами на графике Circos) между двумя сравниваемыми геномами.Порог слияния позволяет объединить две ссылки / ленты, расстояние между которыми находится в пределах определенного пользователем порога, а порог связи позволяет пользователям исключать любые сопоставленные / гомологичные области, размер генома которых меньше заданного пользователем порогового значения. Дорожка гистограммы добавляется во внешнем кольце кругового графика, чтобы указать процент картированных областей, что позволяет пользователям быстро идентифицировать потенциальные отступы (обозначенные белыми промежутками) и области картирования (обозначенные зелеными диаграммами) между двумя выровненными геномами. Реализация конвейера PGC управляется с помощью сценариев Perl. Этот конвейер производит два типа выходных данных: результаты выравнивания NUCmer и высококачественный график Circos (формат SVG). Пользователи могут бесплатно загрузить эти результаты для публикации или дальнейшего анализа на странице результатов PGC.
Существующий инструмент сравнения генома микробов (MGC) использует метод вычитания генома in silico для идентификации генетических элементов, специфичных для группы штаммов [16]. В то время как инструмент PGC использует файлы генома и NUCmer для выполнения попарного выравнивания генома, инструмент MGC использует in silico фрагментированные последовательности генома и выполняет BLASTN по группам запросов.Напротив, браузер VISTA, хорошо известный своим биологическим применением, может выполнять предварительно вычисленное попарное и множественное выравнивание генома с использованием как глобального, так и локального выравнивания [17]. В отличие от круговых графиков и гистограмм, которые генерируются инструментом PGC, результаты выравнивания, сгенерированные VISTA Browser, отображаются с использованием трека VISTA в формате графика, чтобы показать сохраненные области. Кроме того, основанный на Java инструмент сравнения Artemis (ACT) с открытым исходным кодом требует, чтобы пользователи сгенерировали файл сравнения, который идентифицирует области гомологии между сборочным и эталонным геномом с помощью таких программ, как BLASTN, TBLASTX или Mummer, которые должны быть загружены в ACT [18].Сравнительная визуализация ACT выполняется с использованием компонентов Artemis. Напротив, наш инструмент PGC обеспечивает однопоточный процесс попарного выравнивания генома и мгновенное отображение графика Circos сравнительного выравнивания.
Чтобы продемонстрировать полезность PGC, мы сравнили S . mitis B6 (полный геном) и 17/34 (черновой вариант генома) в качестве примера в.
Парное сравнение генома между S . mitis B6 и S . mitis 17/34 с использованием инструмента PGC, встроенного в StreptoBase.
50% идентичности последовательностей и 50% покрытия последовательностей использовали для сравнения штаммов с использованием инструмента PGC. A и B выделяют отступы парного сравнения генома между S . mitis B6 и S . mitis 17/34.
Параметры были установлены как 80% от минимального процента, значение по умолчанию — порог связи 1000 бит и порог слияния 2000 бит. S . mitis B6 был выделен в Германии, а S . mitis 17/34 был выделен из уретры российского пациента с уретритом.На основе сгенерированного графика PGC оба значения S . mitis геномов обычно имели высокое сходство, так как большинство их геномных областей могли быть выровнены (). Одной из особенностей графика PGC является его способность быстро определять предполагаемые отступы посредством визуализации пробелов в графике, что подтверждается информацией, отображаемой на дорожке гистограммы. Например, два разрыва () указывают на отсутствие геномных областей в S . mitis 17/34 геном.Внешняя круглая полоса графика показывает измерения размера генома, которые составляют примерно 2 МБ для обоих S . mitis геномов. Исходя из разрыва, наблюдаемого в (indel «A»), потеря гена произошла около позиции 400 000 п.н.
Затем мы исследовали гены, расположенные в индексе «A» в S . mitis B6 () путем визуализации этой области с помощью SGB. Мы идентифицировали множество генов, связанных с фагами, связанных с этой областью. Для дальнейшего изучения этой области мы использовали PHAST (инструмент поиска PHAge) для аннотирования и идентификации последовательностей профагов, обнаруженных в пределах S . mitis B6 геном (You Zhou et al., 2011). Интактный профаг размером 56 КБ с 82 CDS и содержанием GC 39,9% был обнаружен с 390 924 до 446 969 пар оснований. Начиная с S . mitis B6 — это полный геном, поэтому мы можем указать положение пары оснований непосредственно в нашем файле аннотации B6. Согласно результатам PHAST, это интактный профаг S . mitis B6 содержал фаг-ассоциированные гены, включая белок фаговой интегразы, фаговый CI-подобный репрессор, фаг-связывающий белок, портальный белок фага, белок морфогенеза головки фага семейства SPP1 и белки фагового капсида. Поэтому мы предлагаем S . mitis B6, возможно, недавно приобрел этот интактный профаг. Графическое изображение интактного профага с различными типами фаг-связанных генов показано на.
В S обнаружен интактный профаг. митис B6. Этот профаг имеет 85 предсказанных генов.
На основании инделя «B», обнаруженного на графике PGC, мы выявили неполный профаг размером 24 Кбайт с содержанием GC 39,17%, расположенный в положениях от 1356040 пар оснований до 1380128 пар оснований. Интересно, что эта область содержит полный оперон атр , регулируемый Белок CcpA в этом неполном профаге S . mitis геном B6. Гены оперона atp показаны в. Эти гены, кодирующие АТФ-синтазы, обычно используются оральными стрептококками для адаптации к кислой среде хозяина за счет создания более щелочной внутренней системы.
Таблица 5
Синтазы АТФ в опероне atp из S . митис B6.
Имя Локус Метка
Джина Функциональных аннотации
smi_1315
atpE
АТФ-синтазы С-цепью (ЕС 3.6.3.14)
smi_1314
atpB
Цепь АТФ-синтазы А (EC 3.6.3.14)
smi_1313
atpF-синтаза Цепь B 901 3.14)
smi_1312
atpH
Дельта-цепь АТФ-синтазы (EC 3. 6.3.14)
smi_1311
в цепи синтаза альфа-6.3.14)
smi_1310
atpG
Гамма-цепь АТФ-синтазы (EC 3.6.3.14)
smi_1309
beta. 3.14.Следовательно, возможно, что приобретение этого оперона atp осуществляется неполным профагом S . mitis B6 посредством горизонтального переноса генов помог его комменсальному статусу поддерживать оптимальный уровень pH для биоэнергетических процессов S . mitis клеток B6.
PathoProT был разработан для прогнозирования генов вирулентности путем сравнения аминокислотных последовательностей Streptococcus с базой данных факторов вирулентности (VFDB) [20]. PathoProT использует автономные инструменты BLAST, загруженные с веб-сайта NCBI. VFDB (версия 2012) в настоящее время содержит набор из 19775 экспериментально подтвержденных генов вирулентности, происходящих от широкого круга различных видов бактерий, что обеспечивает полезный ресурс для поиска гомологии последовательностей. Пользователи могут выбрать список из штаммов Streptococcus для сравнительного анализа и установить пороговое значение, например, идентичность генома и полноту для поиска BLAST через нашу предоставленную онлайн-форму.Параметры по умолчанию для конвейера PathoProT установлены на 50% идентичности последовательностей и 50% полноты последовательности для поиска и идентификации генов ортологичной вирулентности в выбранных геномах Streptococcus . Однако пользователи могут применять желаемые отсечки для поиска гомологии, чтобы достичь оптимальных уровней строгости в своих анализах.
Вкратце, конвейер PathoProT в основном реализован с использованием Perl. Собственные сценарии Perl будут обрабатывать выходные данные BLAST (сгенерированные путем поиска этих последовательностей запросов в VFDB) для каждого предсказанного RAST белка (последовательности запроса) в выбранных пользователем геномах и определять предполагаемую вирулентность на основе заданных пользователем параметров.Отфильтрованные результаты BLAST объединяются и организуются в матричную таблицу, содержащую информацию о наличии или отсутствии генов вирулентности (строки) и названия штаммов Streptococcus (столбцы). Наконец, PathoProT передаст и обработает этот вывод с помощью наших собственных сценариев R для иерархической кластеризации (алгоритм полной связи) и создания тепловой карты для визуализации. Штаммы Streptococcus будут отсортированы на основе их профилей генов вирулентности (), и будет построено филогенетическое дерево, пользователи смогут оценить взаимосвязи между близкородственными S . mitis групповых видов / штаммов, а также соответствующие им гены вирулентности образуют заметные кластеры на дендрограммах. Таким образом, этот сравнительный анализ патогеномики может дать отличное представление о профилях генов вирулентности у разных видов Streptococcus . Например, не существует инструмента биоинформатики, который выполняет те же функции, что и PathoProT, а именно прогнозирует и позволяет сравнивать гены вирулентности в геномах разных видов бактерий.
Блок-схема PathoProT.
PathoProT в основном реализован с использованием сценариев Perl и R. Входными данными PathoProT будут списки генов для выбранных штаммов / геномов, и конвейер будет генерировать тепловую карту в конце процесса.
Чтобы продемонстрировать особенности или функциональные возможности PathoProT, мы представляем сравнительное исследование патогеномики среди S . mitis группируют бактерии, используя порог в 50% как для идентичности последовательностей, так и для покрытия, чтобы получить представление об их профилях генов вирулентности.На основании созданной тепловой карты PathoProT ряд предполагаемых генов вирулентности, по-видимому, сохраняется среди всех видов группы мититов (). Консервативные гены hasC (hasC1 или SMU . 322c) , которые кодируют UTP-глюкозо-1-фосфат уридилилтрансферазу (или UDP-пирофосфорилазу глюкозы) (M6Spy1871), участвуют в синтезе гиалуроновой кислоты вдоль (HA) капсулы. с двумя соседними генами: hasA и hasB в пределах имеет оперон.[21]. Фактически, это Streptococcus pneumoniae , наиболее патогенный вид S . Группа mitis обладает полисахаридной капсулой, которая способствует бактериальному патогенезу [22]. В Streptococcus HA обнаружен, поскольку материал капсулы стрептококка у некоторых видов является важным фактором вирулентности, эффективно маскируя бактерии от распознавания иммунной системой хозяина [23,24], а также защищая их от реактивных оксидов, выделяемых лейкоцитами [25] ].Кроме того, возможно, что ГК играет важную роль в прикреплении стрептококков группы мититов и колонизации эпителиальных клеток, что приводит к устойчивости бактерий к фагоцитозу макрофагами [26–28].
Информативная тепловая карта, созданная инструментом PathoProT.
(A) Список консервативных генов вирулентности, переносимых всеми видами группы mitis, и (B) гены, связанные с синтезом RGP, которые могут дифференцировать M Clade и S Clade. Наличие гена вирулентности было отмечено красным цветом, а отсутствие генов вирулентности было отмечено черным.
Другой консервативный ген вирулентности, slrA , кодирует стрептококковую липопротеин ротамазу А, которая является одним из основных поверхностных белков, экспрессируемых S . pneumoniae . Этот ген является важным циклофилином, который модулирует биологическую функцию белков вирулентности на первой стадии пневмококковой инфекции [29]. Вероятно, что ген slrA способствует инвазии клеток-хозяев и облегчает колонизацию пневмококков и присоединение к ним в S . бактерий группы mitis [30,31]. Кроме того, сообщалось, что дефицит slrA снижает вирулентность бактерий из-за его влияния на адгезию и интернализацию эпителиальными и эндотелиальными клетками [29]. Аналогичным образом, консервативный ген lmb кодирует ламинин-связывающий белок, который впервые был идентифицирован в Streptococcus agalactiae [32]. Практически идентичные адгезины позже были обнаружены как у Streptococcus suis [33], так и у Streptococcus pyogenes [34,35].Было предложено, что адгезины lmb помогают в бактериальном патогенезе посредством инвазии поврежденного эпителия [36]. В целом, многие поверхностные липопротеины и адгезины, которые важны для вирулентности и патогенных инфекций, в S очень консервативны. mitis группа бактерий.
Согласно филогенетическому дереву, сгенерированному в левой части тепловой карты PathoProT (), группу мититов можно четко разделить на две категории: S Clade ( S . sanguinis , S . гордонии , S . parasanguinis , S . Австралийский , S . cristatus и S . oligofermentans ) и M Clade ( S . mitis , S . infantis , S . tigurinus , S . oralis и S ). Это филогенетическое родство S . mitis групповых видов указывает на близкое родство между видами в M Clade и между видами в S Clade. Интересно, что мы обнаружили, что гены rgp могут использоваться для дифференциации двух разных кладов на тепловой карте. Например, эти маркерные гены присутствуют у всех видов S Clade, но отсутствуют у всех видов M Clade.
Кластер генов rgp (B, C, D, F и G) отвечает за синтез полисахарида рамнозы-глюкозы (RGP) в Streptococcus mutans .Примечательно, что подобные гены участвуют в синтезе рамнана у Escherichia coli [37]. Фактически, было предложено, чтобы E . coli и S . mutans имеют общий путь синтеза рамнана, основанный на их сходстве в синтезе RGP [37]. Функция rgpB заключается в переносе второго остатка рамнозы на остаток рамнозы на N -ацетилглюкозамине, связанном с липидным носителем, за которым следует rgpF , который позже катализирует перенос третьего остатка рамнозы на вторую рамнозу. остаток полученного гликолипидного носителя.И rgpB , и rgpF предположительно должны работать поочередно при удлинении цепи рамнана. Гомологичные рамнозилтрансферазы rgpB и rgpF были обнаружены у Streptococcus thermophilus (STER1436) и Streptococcus gordonii (SGO1022). С другой стороны, гены rgpC и rgpD кодируют предполагаемые переносчики ABC, специфичные для RGP (гомологичный STER1434 в S . thermophilus и гомологичный SGO1024 в S . gordonii ), которые играют роль в экспорте полисахаридов [37]. Ген rgpG (гомолог S . gordonii SGO1723) инициирует синтез RGP путем переноса N -ацетилглюкозамин-1-фосфата на липидный носитель [38].
Гены rgp также вовлечены в патогенез нескольких видов Streptococcus . Например, rgp играет важную роль в бактериальной вирулентности, а также вызывает воспалительную реакцию в S . suis [39]. Индукция инфекционного эндокардита по S . Сообщалось, что mutans запускается генами rgp посредством высвобождения оксида азота [40], агрегации тромбоцитов [41] и придания устойчивости к фагоцитозу полиморфно-ядерными лейкоцитами человека [42]. Следовательно, S Clade S . Виды группы mitis , которые продуцируют эти богатые рамнозой полимеры, могут демонстрировать патогенез, отличный от видов M Clade Streptococcus , с целью установления большей вирулентности и увеличения выживаемости в клетках-хозяевах.Недавнее исследование выявило группу стрептококков Sanguinis в качестве распространенного возбудителя транзиторной бактериемии, которая потенциально может привести к инфекционному эндокардиту. Сообщалось также, что эта группа присутствовала в нескольких случаях вирулентной сепсисной инфекции у пациентов с нейтропенией [43].
Инструменты поиска последовательности
Мы включили в StreptoBase два типа механизмов BLAST, стандартный BLAST и VFDB BLAST, для поиска штаммов Streptococcus , ближайших к штамму запроса. Эти эксклюзивные поиски BLAST функционально основаны на автономном инструменте BLAST [44], загруженном из NCBI. Оба механизма BLAST поддерживают три типа функций BLAST, а именно BLASTN, BLASTP и BLASTX. Пользователи могут определять полноту генома (%) и идентичность генома (%) в формах отправки инструментов BLAST. Эти специализированные инструменты BLAST призваны облегчить пользователям поиск сходства их запрашиваемых последовательностей с последовательностями генома Streptococcus , последовательностями генов (стандартный BLAST), а также с генами вирулентности VFDB (VFDB BLAST), что позволяет пользователям проверить, действительно ли их представляющие интерес гены являются генами потенциальной вирулентности с использованием подхода гомологии последовательностей.
Дальнейшая работа и заключение
С развитием технологии NGS будет секвенировано еще видов или штаммов Streptococcus , что создает острую потребность в хранении, просмотре, извлечении и анализе огромных объемов данных генома и разработке специализированных инструментов для сравнительный анализ этих геномов.
Здесь мы успешно описали и продемонстрировали функциональные возможности StreptoBase, в частности разработанные нами биоинформатические конвейеры для анализа геномных данных Streptococcus .
Эта специализированная биологическая база данных будет постоянно обновляться, чтобы предоставлять последние обновления генома и исследования, связанные с родом Streptococcus , а также обеспечивать точность и полезность S . mitis данные генома вида группы и аннотация. Мы ожидаем, что StreptoBase будет служить полезным ресурсом и платформой для анализа, особенно для сравнительного анализа S . mitis групповых геномов для исследовательских сообществ.Мы призываем других исследователей или исследовательские группы вносить предложения и делиться с нами своими аннотациями, мнениями и собранными данными по адресу ym.ude.mu@grig.
Группа геномных ресурсов и анализа Oral Streptococcus mitis
Функции базы данных и встроенные инструменты биоинформатики
S . mitis Виды группы являются важными колонизаторами ротовой полости и иногда связаны с серьезными инфекциями [15]. Кроме того, недавно было высказано предположение, что эти организмы играют важную роль в патогенезе гриппа [8].Таким образом, геномное исследование разнообразных S . mitis Бактерии группы необходимы для понимания того, как эти микроорганизмы переходят от комменсального образа жизни во рту к последующему патогенезу. Однако для широкого спектра S не существует специализированной базы данных генома. mitis групповых геномов для сравнительной геномики. Хотя большинство баз данных по биологическим геномам сосредоточены только на содержании генома и генетической изменчивости, мы определили необходимость создания функциональных биоинформатических инструментов для исследования детерминант вирулентности в геномах посредством сравнительной патогеномики, а также для сравнения содержания генома и генетической изменчивости в пределах S . mitis группа бактерий.
Инструмент парного сравнения генома (PGC)
Мы разработали и настроили веб-инструмент PGC для S . mitis группирует бактерии, что позволяет пользователям выбирать и проводить попарные сравнения между двумя выбранными пользователем геномами Streptococcus . Список геномов Streptococcus доступен в инструменте PGC StreptoBase, что позволяет пользователям выбрать два генома Streptococcus для сравнения штаммов или видов.Кроме того, пользователи могут загрузить свои собственные последовательности генома, нуклеотидов или белков, и сравнить их с геномами Streptococcus в StreptoBase.
Вкратце, конвейер PGC поддерживается NUCmer, который разработан для выравнивания полногеномных последовательностей, и Circos, который является хорошо зарекомендовавшим себя инструментом для визуализации генома. После того, как пользователи отправят свои задания на наш сервер, PGC вызовет программу NUCmer для согласования выбранных пользователем геномов, а внутренние сценарии будут использоваться для обработки выходных данных выравнивания генома и создания входных файлов, анализируемых в Circos, чтобы сгенерировать макет круговой идеограммы выравнивания. В отличие от обычного линейного отображения выравниваний, круговая компоновка показывает взаимосвязь между парами позиций с кариотипами и связями, кодирующими положение, размер и ориентацию связанных геномных элементов.
В веб-интерфейсе PGC предусмотрены три определяемых пользователем параметра, включая минимальный процент идентичности (%), порог слияния (bp) и порог связи (bp). Граница минимального процента идентичности определяет гомологичную область (представленную связями / лентами на графике Circos) между двумя сравниваемыми геномами.Порог слияния позволяет объединить две ссылки / ленты, расстояние между которыми находится в пределах определенного пользователем порога, а порог связи позволяет пользователям исключать любые сопоставленные / гомологичные области, размер генома которых меньше заданного пользователем порогового значения. Дорожка гистограммы добавляется во внешнем кольце кругового графика, чтобы указать процент картированных областей, что позволяет пользователям быстро идентифицировать потенциальные отступы (обозначенные белыми промежутками) и области картирования (обозначенные зелеными диаграммами) между двумя выровненными геномами. Реализация конвейера PGC управляется с помощью сценариев Perl. Этот конвейер производит два типа выходных данных: результаты выравнивания NUCmer и высококачественный график Circos (формат SVG). Пользователи могут бесплатно загрузить эти результаты для публикации или дальнейшего анализа на странице результатов PGC.
Существующий инструмент сравнения генома микробов (MGC) использует метод вычитания генома in silico для идентификации генетических элементов, специфичных для группы штаммов [16]. В то время как инструмент PGC использует файлы генома и NUCmer для выполнения попарного выравнивания генома, инструмент MGC использует in silico фрагментированные последовательности генома и выполняет BLASTN по группам запросов.Напротив, браузер VISTA, хорошо известный своим биологическим применением, может выполнять предварительно вычисленное попарное и множественное выравнивание генома с использованием как глобального, так и локального выравнивания [17]. В отличие от круговых графиков и гистограмм, которые генерируются инструментом PGC, результаты выравнивания, сгенерированные VISTA Browser, отображаются с использованием трека VISTA в формате графика, чтобы показать сохраненные области. Кроме того, основанный на Java инструмент сравнения Artemis (ACT) с открытым исходным кодом требует, чтобы пользователи сгенерировали файл сравнения, который идентифицирует области гомологии между сборочным и эталонным геномом с помощью таких программ, как BLASTN, TBLASTX или Mummer, которые должны быть загружены в ACT [18].Сравнительная визуализация ACT выполняется с использованием компонентов Artemis. Напротив, наш инструмент PGC обеспечивает однопоточный процесс попарного выравнивания генома и мгновенное отображение графика Circos сравнительного выравнивания.
Чтобы продемонстрировать полезность PGC, мы сравнили S . mitis B6 (полный геном) и 17/34 (черновой вариант генома) в качестве примера в.
Парное сравнение генома между S . mitis B6 и S . mitis 17/34 с использованием инструмента PGC, встроенного в StreptoBase.
50% идентичности последовательностей и 50% покрытия последовательностей использовали для сравнения штаммов с использованием инструмента PGC. A и B выделяют отступы парного сравнения генома между S . mitis B6 и S . mitis 17/34.
Параметры были установлены как 80% от минимального процента, значение по умолчанию — порог связи 1000 бит и порог слияния 2000 бит. S . mitis B6 был выделен в Германии, а S . mitis 17/34 был выделен из уретры российского пациента с уретритом.На основе сгенерированного графика PGC оба значения S . mitis геномов обычно имели высокое сходство, так как большинство их геномных областей могли быть выровнены (). Одной из особенностей графика PGC является его способность быстро определять предполагаемые отступы посредством визуализации пробелов в графике, что подтверждается информацией, отображаемой на дорожке гистограммы. Например, два разрыва () указывают на отсутствие геномных областей в S . mitis 17/34 геном.Внешняя круглая полоса графика показывает измерения размера генома, которые составляют примерно 2 МБ для обоих S . mitis геномов. Исходя из разрыва, наблюдаемого в (indel «A»), потеря гена произошла около позиции 400 000 п.н.
Затем мы исследовали гены, расположенные в индексе «A» в S . mitis B6 () путем визуализации этой области с помощью SGB. Мы идентифицировали множество генов, связанных с фагами, связанных с этой областью. Для дальнейшего изучения этой области мы использовали PHAST (инструмент поиска PHAge) для аннотирования и идентификации последовательностей профагов, обнаруженных в пределах S . mitis B6 геном (You Zhou et al., 2011). Интактный профаг размером 56 КБ с 82 CDS и содержанием GC 39,9% был обнаружен с 390 924 до 446 969 пар оснований. Начиная с S . mitis B6 — это полный геном, поэтому мы можем указать положение пары оснований непосредственно в нашем файле аннотации B6. Согласно результатам PHAST, это интактный профаг S . mitis B6 содержал фаг-ассоциированные гены, включая белок фаговой интегразы, фаговый CI-подобный репрессор, фаг-связывающий белок, портальный белок фага, белок морфогенеза головки фага семейства SPP1 и белки фагового капсида. Поэтому мы предлагаем S . mitis B6, возможно, недавно приобрел этот интактный профаг. Графическое изображение интактного профага с различными типами фаг-связанных генов показано на.
В S обнаружен интактный профаг. митис B6. Этот профаг имеет 85 предсказанных генов.
На основании инделя «B», обнаруженного на графике PGC, мы выявили неполный профаг размером 24 Кбайт с содержанием GC 39,17%, расположенный в положениях от 1356040 пар оснований до 1380128 пар оснований. Интересно, что эта область содержит полный оперон атр , регулируемый Белок CcpA в этом неполном профаге S . mitis геном B6. Гены оперона atp показаны в. Эти гены, кодирующие АТФ-синтазы, обычно используются оральными стрептококками для адаптации к кислой среде хозяина за счет создания более щелочной внутренней системы.
Таблица 5
Синтазы АТФ в опероне atp из S . митис B6.
Метка локуса
Имя гена
Функциональная аннотация
smi_1315
atpE
АТФ-синтаза C-цепочка (EC 3.6.3.14)
smi_1314
atpB
Цепь АТФ-синтазы А (EC 3.6.3.14)
smi_1313
atpF-синтаза Цепь B 901 3.14)
smi_1312
atpH
Дельта-цепь АТФ-синтазы (EC 3. 6.3.14)
smi_1311
в цепи синтаза альфа-6.3.14)
smi_1310
atpG
Гамма-цепь АТФ-синтазы (EC 3.6.3.14)
smi_1309
beta. 3.14.Следовательно, возможно, что приобретение этого оперона atp осуществляется неполным профагом S . mitis B6 посредством горизонтального переноса генов помог его комменсальному статусу поддерживать оптимальный уровень pH для биоэнергетических процессов S . mitis клеток B6.
PathoProT был разработан для прогнозирования генов вирулентности путем сравнения аминокислотных последовательностей Streptococcus с базой данных факторов вирулентности (VFDB) [20]. PathoProT использует автономные инструменты BLAST, загруженные с веб-сайта NCBI. VFDB (версия 2012) в настоящее время содержит набор из 19775 экспериментально подтвержденных генов вирулентности, происходящих от широкого круга различных видов бактерий, что обеспечивает полезный ресурс для поиска гомологии последовательностей. Пользователи могут выбрать список из штаммов Streptococcus для сравнительного анализа и установить пороговое значение, например, идентичность генома и полноту для поиска BLAST через нашу предоставленную онлайн-форму.Параметры по умолчанию для конвейера PathoProT установлены на 50% идентичности последовательностей и 50% полноты последовательности для поиска и идентификации генов ортологичной вирулентности в выбранных геномах Streptococcus . Однако пользователи могут применять желаемые отсечки для поиска гомологии, чтобы достичь оптимальных уровней строгости в своих анализах.
Вкратце, конвейер PathoProT в основном реализован с использованием Perl. Собственные сценарии Perl будут обрабатывать выходные данные BLAST (сгенерированные путем поиска этих последовательностей запросов в VFDB) для каждого предсказанного RAST белка (последовательности запроса) в выбранных пользователем геномах и определять предполагаемую вирулентность на основе заданных пользователем параметров.Отфильтрованные результаты BLAST объединяются и организуются в матричную таблицу, содержащую информацию о наличии или отсутствии генов вирулентности (строки) и названия штаммов Streptococcus (столбцы). Наконец, PathoProT передаст и обработает этот вывод с помощью наших собственных сценариев R для иерархической кластеризации (алгоритм полной связи) и создания тепловой карты для визуализации. Штаммы Streptococcus будут отсортированы на основе их профилей генов вирулентности (), и будет построено филогенетическое дерево, пользователи смогут оценить взаимосвязи между близкородственными S . mitis групповых видов / штаммов, а также соответствующие им гены вирулентности образуют заметные кластеры на дендрограммах. Таким образом, этот сравнительный анализ патогеномики может дать отличное представление о профилях генов вирулентности у разных видов Streptococcus . Например, не существует инструмента биоинформатики, который выполняет те же функции, что и PathoProT, а именно прогнозирует и позволяет сравнивать гены вирулентности в геномах разных видов бактерий.
Блок-схема PathoProT.
PathoProT в основном реализован с использованием сценариев Perl и R. Входными данными PathoProT будут списки генов для выбранных штаммов / геномов, и конвейер будет генерировать тепловую карту в конце процесса.
Чтобы продемонстрировать особенности или функциональные возможности PathoProT, мы представляем сравнительное исследование патогеномики среди S . mitis группируют бактерии, используя порог в 50% как для идентичности последовательностей, так и для покрытия, чтобы получить представление об их профилях генов вирулентности.На основании созданной тепловой карты PathoProT ряд предполагаемых генов вирулентности, по-видимому, сохраняется среди всех видов группы мититов (). Консервативные гены hasC (hasC1 или SMU . 322c) , которые кодируют UTP-глюкозо-1-фосфат уридилилтрансферазу (или UDP-пирофосфорилазу глюкозы) (M6Spy1871), участвуют в синтезе гиалуроновой кислоты вдоль (HA) капсулы. с двумя соседними генами: hasA и hasB в пределах имеет оперон.[21]. Фактически, это Streptococcus pneumoniae , наиболее патогенный вид S . Группа mitis обладает полисахаридной капсулой, которая способствует бактериальному патогенезу [22]. В Streptococcus HA обнаружен, поскольку материал капсулы стрептококка у некоторых видов является важным фактором вирулентности, эффективно маскируя бактерии от распознавания иммунной системой хозяина [23,24], а также защищая их от реактивных оксидов, выделяемых лейкоцитами [25] ].Кроме того, возможно, что ГК играет важную роль в прикреплении стрептококков группы мититов и колонизации эпителиальных клеток, что приводит к устойчивости бактерий к фагоцитозу макрофагами [26–28].
Информативная тепловая карта, созданная инструментом PathoProT.
(A) Список консервативных генов вирулентности, переносимых всеми видами группы mitis, и (B) гены, связанные с синтезом RGP, которые могут дифференцировать M Clade и S Clade. Наличие гена вирулентности было отмечено красным цветом, а отсутствие генов вирулентности было отмечено черным.
Другой консервативный ген вирулентности, slrA , кодирует стрептококковую липопротеин ротамазу А, которая является одним из основных поверхностных белков, экспрессируемых S . pneumoniae . Этот ген является важным циклофилином, который модулирует биологическую функцию белков вирулентности на первой стадии пневмококковой инфекции [29]. Вероятно, что ген slrA способствует инвазии клеток-хозяев и облегчает колонизацию пневмококков и присоединение к ним в S . бактерий группы mitis [30,31]. Кроме того, сообщалось, что дефицит slrA снижает вирулентность бактерий из-за его влияния на адгезию и интернализацию эпителиальными и эндотелиальными клетками [29]. Аналогичным образом, консервативный ген lmb кодирует ламинин-связывающий белок, который впервые был идентифицирован в Streptococcus agalactiae [32]. Практически идентичные адгезины позже были обнаружены как у Streptococcus suis [33], так и у Streptococcus pyogenes [34,35].Было предложено, что адгезины lmb помогают в бактериальном патогенезе посредством инвазии поврежденного эпителия [36]. В целом, многие поверхностные липопротеины и адгезины, которые важны для вирулентности и патогенных инфекций, в S очень консервативны. mitis группа бактерий.
Согласно филогенетическому дереву, сгенерированному в левой части тепловой карты PathoProT (), группу мититов можно четко разделить на две категории: S Clade ( S . sanguinis , S . гордонии , S . parasanguinis , S . Австралийский , S . cristatus и S . oligofermentans ) и M Clade ( S . mitis , S . infantis , S . tigurinus , S . oralis и S ). Это филогенетическое родство S . mitis групповых видов указывает на близкое родство между видами в M Clade и между видами в S Clade. Интересно, что мы обнаружили, что гены rgp могут использоваться для дифференциации двух разных кладов на тепловой карте. Например, эти маркерные гены присутствуют у всех видов S Clade, но отсутствуют у всех видов M Clade.
Кластер генов rgp (B, C, D, F и G) отвечает за синтез полисахарида рамнозы-глюкозы (RGP) в Streptococcus mutans .Примечательно, что подобные гены участвуют в синтезе рамнана у Escherichia coli [37]. Фактически, было предложено, чтобы E . coli и S . mutans имеют общий путь синтеза рамнана, основанный на их сходстве в синтезе RGP [37]. Функция rgpB заключается в переносе второго остатка рамнозы на остаток рамнозы на N -ацетилглюкозамине, связанном с липидным носителем, за которым следует rgpF , который позже катализирует перенос третьего остатка рамнозы на вторую рамнозу. остаток полученного гликолипидного носителя.И rgpB , и rgpF предположительно должны работать поочередно при удлинении цепи рамнана. Гомологичные рамнозилтрансферазы rgpB и rgpF были обнаружены у Streptococcus thermophilus (STER1436) и Streptococcus gordonii (SGO1022). С другой стороны, гены rgpC и rgpD кодируют предполагаемые переносчики ABC, специфичные для RGP (гомологичный STER1434 в S . thermophilus и гомологичный SGO1024 в S . gordonii ), которые играют роль в экспорте полисахаридов [37]. Ген rgpG (гомолог S . gordonii SGO1723) инициирует синтез RGP путем переноса N -ацетилглюкозамин-1-фосфата на липидный носитель [38].
Гены rgp также вовлечены в патогенез нескольких видов Streptococcus . Например, rgp играет важную роль в бактериальной вирулентности, а также вызывает воспалительную реакцию в S . suis [39]. Индукция инфекционного эндокардита по S . Сообщалось, что mutans запускается генами rgp посредством высвобождения оксида азота [40], агрегации тромбоцитов [41] и придания устойчивости к фагоцитозу полиморфно-ядерными лейкоцитами человека [42]. Следовательно, S Clade S . Виды группы mitis , которые продуцируют эти богатые рамнозой полимеры, могут демонстрировать патогенез, отличный от видов M Clade Streptococcus , с целью установления большей вирулентности и увеличения выживаемости в клетках-хозяевах.Недавнее исследование выявило группу стрептококков Sanguinis в качестве распространенного возбудителя транзиторной бактериемии, которая потенциально может привести к инфекционному эндокардиту. Сообщалось также, что эта группа присутствовала в нескольких случаях вирулентной сепсисной инфекции у пациентов с нейтропенией [43].
Инструменты поиска последовательности
Мы включили в StreptoBase два типа механизмов BLAST, стандартный BLAST и VFDB BLAST, для поиска штаммов Streptococcus , ближайших к штамму запроса. Эти эксклюзивные поиски BLAST функционально основаны на автономном инструменте BLAST [44], загруженном из NCBI. Оба механизма BLAST поддерживают три типа функций BLAST, а именно BLASTN, BLASTP и BLASTX. Пользователи могут определять полноту генома (%) и идентичность генома (%) в формах отправки инструментов BLAST. Эти специализированные инструменты BLAST призваны облегчить пользователям поиск сходства их запрашиваемых последовательностей с последовательностями генома Streptococcus , последовательностями генов (стандартный BLAST), а также с генами вирулентности VFDB (VFDB BLAST), что позволяет пользователям проверить, действительно ли их представляющие интерес гены являются генами потенциальной вирулентности с использованием подхода гомологии последовательностей.
Дальнейшая работа и заключение
С развитием технологии NGS будет секвенировано еще видов или штаммов Streptococcus , что создает острую потребность в хранении, просмотре, извлечении и анализе огромных объемов данных генома и разработке специализированных инструментов для сравнительный анализ этих геномов.
Здесь мы успешно описали и продемонстрировали функциональные возможности StreptoBase, в частности разработанные нами биоинформатические конвейеры для анализа геномных данных Streptococcus .
Эта специализированная биологическая база данных будет постоянно обновляться, чтобы предоставлять последние обновления генома и исследования, связанные с родом Streptococcus , а также обеспечивать точность и полезность S . mitis данные генома вида группы и аннотация. Мы ожидаем, что StreptoBase будет служить полезным ресурсом и платформой для анализа, особенно для сравнительного анализа S . mitis групповых геномов для исследовательских сообществ.Мы призываем других исследователей или исследовательские группы вносить предложения и делиться с нами своими аннотациями, мнениями и собранными данными по адресу ym.ude.mu@grig.
Группа геномных ресурсов и анализа Oral Streptococcus mitis
Функции базы данных и встроенные инструменты биоинформатики
S . mitis Виды группы являются важными колонизаторами ротовой полости и иногда связаны с серьезными инфекциями [15]. Кроме того, недавно было высказано предположение, что эти организмы играют важную роль в патогенезе гриппа [8].Таким образом, геномное исследование разнообразных S . mitis Бактерии группы необходимы для понимания того, как эти микроорганизмы переходят от комменсального образа жизни во рту к последующему патогенезу. Однако для широкого спектра S не существует специализированной базы данных генома. mitis групповых геномов для сравнительной геномики. Хотя большинство баз данных по биологическим геномам сосредоточены только на содержании генома и генетической изменчивости, мы определили необходимость создания функциональных биоинформатических инструментов для исследования детерминант вирулентности в геномах посредством сравнительной патогеномики, а также для сравнения содержания генома и генетической изменчивости в пределах S . mitis группа бактерий.
Инструмент парного сравнения генома (PGC)
Мы разработали и настроили веб-инструмент PGC для S . mitis группирует бактерии, что позволяет пользователям выбирать и проводить попарные сравнения между двумя выбранными пользователем геномами Streptococcus . Список геномов Streptococcus доступен в инструменте PGC StreptoBase, что позволяет пользователям выбрать два генома Streptococcus для сравнения штаммов или видов.Кроме того, пользователи могут загрузить свои собственные последовательности генома, нуклеотидов или белков, и сравнить их с геномами Streptococcus в StreptoBase.
Вкратце, конвейер PGC поддерживается NUCmer, который разработан для выравнивания полногеномных последовательностей, и Circos, который является хорошо зарекомендовавшим себя инструментом для визуализации генома. После того, как пользователи отправят свои задания на наш сервер, PGC вызовет программу NUCmer для согласования выбранных пользователем геномов, а внутренние сценарии будут использоваться для обработки выходных данных выравнивания генома и создания входных файлов, анализируемых в Circos, чтобы сгенерировать макет круговой идеограммы выравнивания. В отличие от обычного линейного отображения выравниваний, круговая компоновка показывает взаимосвязь между парами позиций с кариотипами и связями, кодирующими положение, размер и ориентацию связанных геномных элементов.
В веб-интерфейсе PGC предусмотрены три определяемых пользователем параметра, включая минимальный процент идентичности (%), порог слияния (bp) и порог связи (bp). Граница минимального процента идентичности определяет гомологичную область (представленную связями / лентами на графике Circos) между двумя сравниваемыми геномами.Порог слияния позволяет объединить две ссылки / ленты, расстояние между которыми находится в пределах определенного пользователем порога, а порог связи позволяет пользователям исключать любые сопоставленные / гомологичные области, размер генома которых меньше заданного пользователем порогового значения. Дорожка гистограммы добавляется во внешнем кольце кругового графика, чтобы указать процент картированных областей, что позволяет пользователям быстро идентифицировать потенциальные отступы (обозначенные белыми промежутками) и области картирования (обозначенные зелеными диаграммами) между двумя выровненными геномами. Реализация конвейера PGC управляется с помощью сценариев Perl. Этот конвейер производит два типа выходных данных: результаты выравнивания NUCmer и высококачественный график Circos (формат SVG). Пользователи могут бесплатно загрузить эти результаты для публикации или дальнейшего анализа на странице результатов PGC.
Существующий инструмент сравнения генома микробов (MGC) использует метод вычитания генома in silico для идентификации генетических элементов, специфичных для группы штаммов [16]. В то время как инструмент PGC использует файлы генома и NUCmer для выполнения попарного выравнивания генома, инструмент MGC использует in silico фрагментированные последовательности генома и выполняет BLASTN по группам запросов.Напротив, браузер VISTA, хорошо известный своим биологическим применением, может выполнять предварительно вычисленное попарное и множественное выравнивание генома с использованием как глобального, так и локального выравнивания [17]. В отличие от круговых графиков и гистограмм, которые генерируются инструментом PGC, результаты выравнивания, сгенерированные VISTA Browser, отображаются с использованием трека VISTA в формате графика, чтобы показать сохраненные области. Кроме того, основанный на Java инструмент сравнения Artemis (ACT) с открытым исходным кодом требует, чтобы пользователи сгенерировали файл сравнения, который идентифицирует области гомологии между сборочным и эталонным геномом с помощью таких программ, как BLASTN, TBLASTX или Mummer, которые должны быть загружены в ACT [18].Сравнительная визуализация ACT выполняется с использованием компонентов Artemis. Напротив, наш инструмент PGC обеспечивает однопоточный процесс попарного выравнивания генома и мгновенное отображение графика Circos сравнительного выравнивания.
Чтобы продемонстрировать полезность PGC, мы сравнили S . mitis B6 (полный геном) и 17/34 (черновой вариант генома) в качестве примера в.
Парное сравнение генома между S . mitis B6 и S . mitis 17/34 с использованием инструмента PGC, встроенного в StreptoBase.
50% идентичности последовательностей и 50% покрытия последовательностей использовали для сравнения штаммов с использованием инструмента PGC. A и B выделяют отступы парного сравнения генома между S . mitis B6 и S . mitis 17/34.
Параметры были установлены как 80% от минимального процента, значение по умолчанию — порог связи 1000 бит и порог слияния 2000 бит. S . mitis B6 был выделен в Германии, а S . mitis 17/34 был выделен из уретры российского пациента с уретритом.На основе сгенерированного графика PGC оба значения S . mitis геномов обычно имели высокое сходство, так как большинство их геномных областей могли быть выровнены (). Одной из особенностей графика PGC является его способность быстро определять предполагаемые отступы посредством визуализации пробелов в графике, что подтверждается информацией, отображаемой на дорожке гистограммы. Например, два разрыва () указывают на отсутствие геномных областей в S . mitis 17/34 геном.Внешняя круглая полоса графика показывает измерения размера генома, которые составляют примерно 2 МБ для обоих S . mitis геномов. Исходя из разрыва, наблюдаемого в (indel «A»), потеря гена произошла около позиции 400 000 п.н.
Затем мы исследовали гены, расположенные в индексе «A» в S . mitis B6 () путем визуализации этой области с помощью SGB. Мы идентифицировали множество генов, связанных с фагами, связанных с этой областью. Для дальнейшего изучения этой области мы использовали PHAST (инструмент поиска PHAge) для аннотирования и идентификации последовательностей профагов, обнаруженных в пределах S . mitis B6 геном (You Zhou et al., 2011). Интактный профаг размером 56 КБ с 82 CDS и содержанием GC 39,9% был обнаружен с 390 924 до 446 969 пар оснований. Начиная с S . mitis B6 — это полный геном, поэтому мы можем указать положение пары оснований непосредственно в нашем файле аннотации B6. Согласно результатам PHAST, это интактный профаг S . mitis B6 содержал фаг-ассоциированные гены, включая белок фаговой интегразы, фаговый CI-подобный репрессор, фаг-связывающий белок, портальный белок фага, белок морфогенеза головки фага семейства SPP1 и белки фагового капсида. Поэтому мы предлагаем S . mitis B6, возможно, недавно приобрел этот интактный профаг. Графическое изображение интактного профага с различными типами фаг-связанных генов показано на.
В S обнаружен интактный профаг. митис B6. Этот профаг имеет 85 предсказанных генов.
На основании инделя «B», обнаруженного на графике PGC, мы выявили неполный профаг размером 24 Кбайт с содержанием GC 39,17%, расположенный в положениях от 1356040 пар оснований до 1380128 пар оснований. Интересно, что эта область содержит полный оперон атр , регулируемый Белок CcpA в этом неполном профаге S . mitis геном B6. Гены оперона atp показаны в. Эти гены, кодирующие АТФ-синтазы, обычно используются оральными стрептококками для адаптации к кислой среде хозяина за счет создания более щелочной внутренней системы.
Таблица 5
Синтазы АТФ в опероне atp из S . митис B6.
Метка локуса
Имя гена
Функциональная аннотация
smi_1315
atpE
АТФ-синтаза C-цепочка (EC 3.6.3.14)
smi_1314
atpB
Цепь АТФ-синтазы А (EC 3.6.3.14)
smi_1313
atpF-синтаза Цепь B 901 3.14)
smi_1312
atpH
Дельта-цепь АТФ-синтазы (EC 3. 6.3.14)
smi_1311
в цепи синтаза альфа-6.3.14)
smi_1310
atpG
Гамма-цепь АТФ-синтазы (EC 3.6.3.14)
smi_1309
beta. 3.14.Следовательно, возможно, что приобретение этого оперона atp осуществляется неполным профагом S . mitis B6 посредством горизонтального переноса генов помог его комменсальному статусу поддерживать оптимальный уровень pH для биоэнергетических процессов S . mitis клеток B6.
PathoProT был разработан для прогнозирования генов вирулентности путем сравнения аминокислотных последовательностей Streptococcus с базой данных факторов вирулентности (VFDB) [20]. PathoProT использует автономные инструменты BLAST, загруженные с веб-сайта NCBI. VFDB (версия 2012) в настоящее время содержит набор из 19775 экспериментально подтвержденных генов вирулентности, происходящих от широкого круга различных видов бактерий, что обеспечивает полезный ресурс для поиска гомологии последовательностей. Пользователи могут выбрать список из штаммов Streptococcus для сравнительного анализа и установить пороговое значение, например, идентичность генома и полноту для поиска BLAST через нашу предоставленную онлайн-форму.Параметры по умолчанию для конвейера PathoProT установлены на 50% идентичности последовательностей и 50% полноты последовательности для поиска и идентификации генов ортологичной вирулентности в выбранных геномах Streptococcus . Однако пользователи могут применять желаемые отсечки для поиска гомологии, чтобы достичь оптимальных уровней строгости в своих анализах.
Вкратце, конвейер PathoProT в основном реализован с использованием Perl. Собственные сценарии Perl будут обрабатывать выходные данные BLAST (сгенерированные путем поиска этих последовательностей запросов в VFDB) для каждого предсказанного RAST белка (последовательности запроса) в выбранных пользователем геномах и определять предполагаемую вирулентность на основе заданных пользователем параметров.Отфильтрованные результаты BLAST объединяются и организуются в матричную таблицу, содержащую информацию о наличии или отсутствии генов вирулентности (строки) и названия штаммов Streptococcus (столбцы). Наконец, PathoProT передаст и обработает этот вывод с помощью наших собственных сценариев R для иерархической кластеризации (алгоритм полной связи) и создания тепловой карты для визуализации. Штаммы Streptococcus будут отсортированы на основе их профилей генов вирулентности (), и будет построено филогенетическое дерево, пользователи смогут оценить взаимосвязи между близкородственными S . mitis групповых видов / штаммов, а также соответствующие им гены вирулентности образуют заметные кластеры на дендрограммах. Таким образом, этот сравнительный анализ патогеномики может дать отличное представление о профилях генов вирулентности у разных видов Streptococcus . Например, не существует инструмента биоинформатики, который выполняет те же функции, что и PathoProT, а именно прогнозирует и позволяет сравнивать гены вирулентности в геномах разных видов бактерий.
Блок-схема PathoProT.
PathoProT в основном реализован с использованием сценариев Perl и R. Входными данными PathoProT будут списки генов для выбранных штаммов / геномов, и конвейер будет генерировать тепловую карту в конце процесса.
Чтобы продемонстрировать особенности или функциональные возможности PathoProT, мы представляем сравнительное исследование патогеномики среди S . mitis группируют бактерии, используя порог в 50% как для идентичности последовательностей, так и для покрытия, чтобы получить представление об их профилях генов вирулентности.На основании созданной тепловой карты PathoProT ряд предполагаемых генов вирулентности, по-видимому, сохраняется среди всех видов группы мититов (). Консервативные гены hasC (hasC1 или SMU . 322c) , которые кодируют UTP-глюкозо-1-фосфат уридилилтрансферазу (или UDP-пирофосфорилазу глюкозы) (M6Spy1871), участвуют в синтезе гиалуроновой кислоты вдоль (HA) капсулы. с двумя соседними генами: hasA и hasB в пределах имеет оперон.[21]. Фактически, это Streptococcus pneumoniae , наиболее патогенный вид S . Группа mitis обладает полисахаридной капсулой, которая способствует бактериальному патогенезу [22]. В Streptococcus HA обнаружен, поскольку материал капсулы стрептококка у некоторых видов является важным фактором вирулентности, эффективно маскируя бактерии от распознавания иммунной системой хозяина [23,24], а также защищая их от реактивных оксидов, выделяемых лейкоцитами [25] ].Кроме того, возможно, что ГК играет важную роль в прикреплении стрептококков группы мититов и колонизации эпителиальных клеток, что приводит к устойчивости бактерий к фагоцитозу макрофагами [26–28].
Информативная тепловая карта, созданная инструментом PathoProT.
(A) Список консервативных генов вирулентности, переносимых всеми видами группы mitis, и (B) гены, связанные с синтезом RGP, которые могут дифференцировать M Clade и S Clade. Наличие гена вирулентности было отмечено красным цветом, а отсутствие генов вирулентности было отмечено черным.
Другой консервативный ген вирулентности, slrA , кодирует стрептококковую липопротеин ротамазу А, которая является одним из основных поверхностных белков, экспрессируемых S . pneumoniae . Этот ген является важным циклофилином, который модулирует биологическую функцию белков вирулентности на первой стадии пневмококковой инфекции [29]. Вероятно, что ген slrA способствует инвазии клеток-хозяев и облегчает колонизацию пневмококков и присоединение к ним в S . бактерий группы mitis [30,31]. Кроме того, сообщалось, что дефицит slrA снижает вирулентность бактерий из-за его влияния на адгезию и интернализацию эпителиальными и эндотелиальными клетками [29]. Аналогичным образом, консервативный ген lmb кодирует ламинин-связывающий белок, который впервые был идентифицирован в Streptococcus agalactiae [32]. Практически идентичные адгезины позже были обнаружены как у Streptococcus suis [33], так и у Streptococcus pyogenes [34,35].Было предложено, что адгезины lmb помогают в бактериальном патогенезе посредством инвазии поврежденного эпителия [36]. В целом, многие поверхностные липопротеины и адгезины, которые важны для вирулентности и патогенных инфекций, в S очень консервативны. mitis группа бактерий.
Согласно филогенетическому дереву, сгенерированному в левой части тепловой карты PathoProT (), группу мититов можно четко разделить на две категории: S Clade ( S . sanguinis , S . гордонии , S . parasanguinis , S . Австралийский , S . cristatus и S . oligofermentans ) и M Clade ( S . mitis , S . infantis , S . tigurinus , S . oralis и S ). Это филогенетическое родство S . mitis групповых видов указывает на близкое родство между видами в M Clade и между видами в S Clade. Интересно, что мы обнаружили, что гены rgp могут использоваться для дифференциации двух разных кладов на тепловой карте. Например, эти маркерные гены присутствуют у всех видов S Clade, но отсутствуют у всех видов M Clade.
Кластер генов rgp (B, C, D, F и G) отвечает за синтез полисахарида рамнозы-глюкозы (RGP) в Streptococcus mutans .Примечательно, что подобные гены участвуют в синтезе рамнана у Escherichia coli [37]. Фактически, было предложено, чтобы E . coli и S . mutans имеют общий путь синтеза рамнана, основанный на их сходстве в синтезе RGP [37]. Функция rgpB заключается в переносе второго остатка рамнозы на остаток рамнозы на N -ацетилглюкозамине, связанном с липидным носителем, за которым следует rgpF , который позже катализирует перенос третьего остатка рамнозы на вторую рамнозу. остаток полученного гликолипидного носителя.И rgpB , и rgpF предположительно должны работать поочередно при удлинении цепи рамнана. Гомологичные рамнозилтрансферазы rgpB и rgpF были обнаружены у Streptococcus thermophilus (STER1436) и Streptococcus gordonii (SGO1022). С другой стороны, гены rgpC и rgpD кодируют предполагаемые переносчики ABC, специфичные для RGP (гомологичный STER1434 в S . thermophilus и гомологичный SGO1024 в S . gordonii ), которые играют роль в экспорте полисахаридов [37]. Ген rgpG (гомолог S . gordonii SGO1723) инициирует синтез RGP путем переноса N -ацетилглюкозамин-1-фосфата на липидный носитель [38].
Гены rgp также вовлечены в патогенез нескольких видов Streptococcus . Например, rgp играет важную роль в бактериальной вирулентности, а также вызывает воспалительную реакцию в S . suis [39]. Индукция инфекционного эндокардита по S . Сообщалось, что mutans запускается генами rgp посредством высвобождения оксида азота [40], агрегации тромбоцитов [41] и придания устойчивости к фагоцитозу полиморфно-ядерными лейкоцитами человека [42]. Следовательно, S Clade S . Виды группы mitis , которые продуцируют эти богатые рамнозой полимеры, могут демонстрировать патогенез, отличный от видов M Clade Streptococcus , с целью установления большей вирулентности и увеличения выживаемости в клетках-хозяевах.Недавнее исследование выявило группу стрептококков Sanguinis в качестве распространенного возбудителя транзиторной бактериемии, которая потенциально может привести к инфекционному эндокардиту. Сообщалось также, что эта группа присутствовала в нескольких случаях вирулентной сепсисной инфекции у пациентов с нейтропенией [43].
Инструменты поиска последовательности
Мы включили в StreptoBase два типа механизмов BLAST, стандартный BLAST и VFDB BLAST, для поиска штаммов Streptococcus , ближайших к штамму запроса. Эти эксклюзивные поиски BLAST функционально основаны на автономном инструменте BLAST [44], загруженном из NCBI. Оба механизма BLAST поддерживают три типа функций BLAST, а именно BLASTN, BLASTP и BLASTX. Пользователи могут определять полноту генома (%) и идентичность генома (%) в формах отправки инструментов BLAST. Эти специализированные инструменты BLAST призваны облегчить пользователям поиск сходства их запрашиваемых последовательностей с последовательностями генома Streptococcus , последовательностями генов (стандартный BLAST), а также с генами вирулентности VFDB (VFDB BLAST), что позволяет пользователям проверить, действительно ли их представляющие интерес гены являются генами потенциальной вирулентности с использованием подхода гомологии последовательностей.
Дальнейшая работа и заключение
С развитием технологии NGS будет секвенировано еще видов или штаммов Streptococcus , что создает острую потребность в хранении, просмотре, извлечении и анализе огромных объемов данных генома и разработке специализированных инструментов для сравнительный анализ этих геномов.
Здесь мы успешно описали и продемонстрировали функциональные возможности StreptoBase, в частности разработанные нами биоинформатические конвейеры для анализа геномных данных Streptococcus .
Эта специализированная биологическая база данных будет постоянно обновляться, чтобы предоставлять последние обновления генома и исследования, связанные с родом Streptococcus , а также обеспечивать точность и полезность S . mitis данные генома вида группы и аннотация. Мы ожидаем, что StreptoBase будет служить полезным ресурсом и платформой для анализа, особенно для сравнительного анализа S . mitis групповых геномов для исследовательских сообществ.Мы призываем других исследователей или исследовательские группы вносить предложения и делиться с нами своими аннотациями, мнениями и собранными данными по адресу ym.ude.mu@grig.
Группа геномных ресурсов и анализа Oral Streptococcus mitis
Функции базы данных и встроенные инструменты биоинформатики
S . mitis Виды группы являются важными колонизаторами ротовой полости и иногда связаны с серьезными инфекциями [15]. Кроме того, недавно было высказано предположение, что эти организмы играют важную роль в патогенезе гриппа [8].Таким образом, геномное исследование разнообразных S . mitis Бактерии группы необходимы для понимания того, как эти микроорганизмы переходят от комменсального образа жизни во рту к последующему патогенезу. Однако для широкого спектра S не существует специализированной базы данных генома. mitis групповых геномов для сравнительной геномики. Хотя большинство баз данных по биологическим геномам сосредоточены только на содержании генома и генетической изменчивости, мы определили необходимость создания функциональных биоинформатических инструментов для исследования детерминант вирулентности в геномах посредством сравнительной патогеномики, а также для сравнения содержания генома и генетической изменчивости в пределах S . mitis группа бактерий.
Инструмент парного сравнения генома (PGC)
Мы разработали и настроили веб-инструмент PGC для S . mitis группирует бактерии, что позволяет пользователям выбирать и проводить попарные сравнения между двумя выбранными пользователем геномами Streptococcus . Список геномов Streptococcus доступен в инструменте PGC StreptoBase, что позволяет пользователям выбрать два генома Streptococcus для сравнения штаммов или видов.Кроме того, пользователи могут загрузить свои собственные последовательности генома, нуклеотидов или белков, и сравнить их с геномами Streptococcus в StreptoBase.
Вкратце, конвейер PGC поддерживается NUCmer, который разработан для выравнивания полногеномных последовательностей, и Circos, который является хорошо зарекомендовавшим себя инструментом для визуализации генома. После того, как пользователи отправят свои задания на наш сервер, PGC вызовет программу NUCmer для согласования выбранных пользователем геномов, а внутренние сценарии будут использоваться для обработки выходных данных выравнивания генома и создания входных файлов, анализируемых в Circos, чтобы сгенерировать макет круговой идеограммы выравнивания. В отличие от обычного линейного отображения выравниваний, круговая компоновка показывает взаимосвязь между парами позиций с кариотипами и связями, кодирующими положение, размер и ориентацию связанных геномных элементов.
В веб-интерфейсе PGC предусмотрены три определяемых пользователем параметра, включая минимальный процент идентичности (%), порог слияния (bp) и порог связи (bp). Граница минимального процента идентичности определяет гомологичную область (представленную связями / лентами на графике Circos) между двумя сравниваемыми геномами.Порог слияния позволяет объединить две ссылки / ленты, расстояние между которыми находится в пределах определенного пользователем порога, а порог связи позволяет пользователям исключать любые сопоставленные / гомологичные области, размер генома которых меньше заданного пользователем порогового значения. Дорожка гистограммы добавляется во внешнем кольце кругового графика, чтобы указать процент картированных областей, что позволяет пользователям быстро идентифицировать потенциальные отступы (обозначенные белыми промежутками) и области картирования (обозначенные зелеными диаграммами) между двумя выровненными геномами. Реализация конвейера PGC управляется с помощью сценариев Perl. Этот конвейер производит два типа выходных данных: результаты выравнивания NUCmer и высококачественный график Circos (формат SVG). Пользователи могут бесплатно загрузить эти результаты для публикации или дальнейшего анализа на странице результатов PGC.
Существующий инструмент сравнения генома микробов (MGC) использует метод вычитания генома in silico для идентификации генетических элементов, специфичных для группы штаммов [16]. В то время как инструмент PGC использует файлы генома и NUCmer для выполнения попарного выравнивания генома, инструмент MGC использует in silico фрагментированные последовательности генома и выполняет BLASTN по группам запросов.Напротив, браузер VISTA, хорошо известный своим биологическим применением, может выполнять предварительно вычисленное попарное и множественное выравнивание генома с использованием как глобального, так и локального выравнивания [17]. В отличие от круговых графиков и гистограмм, которые генерируются инструментом PGC, результаты выравнивания, сгенерированные VISTA Browser, отображаются с использованием трека VISTA в формате графика, чтобы показать сохраненные области. Кроме того, основанный на Java инструмент сравнения Artemis (ACT) с открытым исходным кодом требует, чтобы пользователи сгенерировали файл сравнения, который идентифицирует области гомологии между сборочным и эталонным геномом с помощью таких программ, как BLASTN, TBLASTX или Mummer, которые должны быть загружены в ACT [18].Сравнительная визуализация ACT выполняется с использованием компонентов Artemis. Напротив, наш инструмент PGC обеспечивает однопоточный процесс попарного выравнивания генома и мгновенное отображение графика Circos сравнительного выравнивания.
Чтобы продемонстрировать полезность PGC, мы сравнили S . mitis B6 (полный геном) и 17/34 (черновой вариант генома) в качестве примера в.
Парное сравнение генома между S . mitis B6 и S . mitis 17/34 с использованием инструмента PGC, встроенного в StreptoBase.
50% идентичности последовательностей и 50% покрытия последовательностей использовали для сравнения штаммов с использованием инструмента PGC. A и B выделяют отступы парного сравнения генома между S . mitis B6 и S . mitis 17/34.
Параметры были установлены как 80% от минимального процента, значение по умолчанию — порог связи 1000 бит и порог слияния 2000 бит. S . mitis B6 был выделен в Германии, а S . mitis 17/34 был выделен из уретры российского пациента с уретритом.На основе сгенерированного графика PGC оба значения S . mitis геномов обычно имели высокое сходство, так как большинство их геномных областей могли быть выровнены (). Одной из особенностей графика PGC является его способность быстро определять предполагаемые отступы посредством визуализации пробелов в графике, что подтверждается информацией, отображаемой на дорожке гистограммы. Например, два разрыва () указывают на отсутствие геномных областей в S . mitis 17/34 геном.Внешняя круглая полоса графика показывает измерения размера генома, которые составляют примерно 2 МБ для обоих S . mitis геномов. Исходя из разрыва, наблюдаемого в (indel «A»), потеря гена произошла около позиции 400 000 п.н.
Затем мы исследовали гены, расположенные в индексе «A» в S . mitis B6 () путем визуализации этой области с помощью SGB. Мы идентифицировали множество генов, связанных с фагами, связанных с этой областью. Для дальнейшего изучения этой области мы использовали PHAST (инструмент поиска PHAge) для аннотирования и идентификации последовательностей профагов, обнаруженных в пределах S . mitis B6 геном (You Zhou et al., 2011). Интактный профаг размером 56 КБ с 82 CDS и содержанием GC 39,9% был обнаружен с 390 924 до 446 969 пар оснований. Начиная с S . mitis B6 — это полный геном, поэтому мы можем указать положение пары оснований непосредственно в нашем файле аннотации B6. Согласно результатам PHAST, это интактный профаг S . mitis B6 содержал фаг-ассоциированные гены, включая белок фаговой интегразы, фаговый CI-подобный репрессор, фаг-связывающий белок, портальный белок фага, белок морфогенеза головки фага семейства SPP1 и белки фагового капсида. Поэтому мы предлагаем S . mitis B6, возможно, недавно приобрел этот интактный профаг. Графическое изображение интактного профага с различными типами фаг-связанных генов показано на.
В S обнаружен интактный профаг. митис B6. Этот профаг имеет 85 предсказанных генов.
На основании инделя «B», обнаруженного на графике PGC, мы выявили неполный профаг размером 24 Кбайт с содержанием GC 39,17%, расположенный в положениях от 1356040 пар оснований до 1380128 пар оснований. Интересно, что эта область содержит полный оперон атр , регулируемый Белок CcpA в этом неполном профаге S . mitis геном B6. Гены оперона atp показаны в. Эти гены, кодирующие АТФ-синтазы, обычно используются оральными стрептококками для адаптации к кислой среде хозяина за счет создания более щелочной внутренней системы.
Таблица 5
Синтазы АТФ в опероне atp из S . митис B6.
Метка локуса
Имя гена
Функциональная аннотация
smi_1315
atpE
АТФ-синтаза C-цепочка (EC 3.6.3.14)
smi_1314
atpB
Цепь АТФ-синтазы А (EC 3.6.3.14)
smi_1313
atpF-синтаза Цепь B 901 3.14)
smi_1312
atpH
Дельта-цепь АТФ-синтазы (EC 3. 6.3.14)
smi_1311
в цепи синтаза альфа-6.3.14)
smi_1310
atpG
Гамма-цепь АТФ-синтазы (EC 3.6.3.14)
smi_1309
beta. 3.14.Следовательно, возможно, что приобретение этого оперона atp осуществляется неполным профагом S . mitis B6 посредством горизонтального переноса генов помог его комменсальному статусу поддерживать оптимальный уровень pH для биоэнергетических процессов S . mitis клеток B6.
PathoProT был разработан для прогнозирования генов вирулентности путем сравнения аминокислотных последовательностей Streptococcus с базой данных факторов вирулентности (VFDB) [20]. PathoProT использует автономные инструменты BLAST, загруженные с веб-сайта NCBI. VFDB (версия 2012) в настоящее время содержит набор из 19775 экспериментально подтвержденных генов вирулентности, происходящих от широкого круга различных видов бактерий, что обеспечивает полезный ресурс для поиска гомологии последовательностей. Пользователи могут выбрать список из штаммов Streptococcus для сравнительного анализа и установить пороговое значение, например, идентичность генома и полноту для поиска BLAST через нашу предоставленную онлайн-форму.Параметры по умолчанию для конвейера PathoProT установлены на 50% идентичности последовательностей и 50% полноты последовательности для поиска и идентификации генов ортологичной вирулентности в выбранных геномах Streptococcus . Однако пользователи могут применять желаемые отсечки для поиска гомологии, чтобы достичь оптимальных уровней строгости в своих анализах.
Вкратце, конвейер PathoProT в основном реализован с использованием Perl. Собственные сценарии Perl будут обрабатывать выходные данные BLAST (сгенерированные путем поиска этих последовательностей запросов в VFDB) для каждого предсказанного RAST белка (последовательности запроса) в выбранных пользователем геномах и определять предполагаемую вирулентность на основе заданных пользователем параметров.Отфильтрованные результаты BLAST объединяются и организуются в матричную таблицу, содержащую информацию о наличии или отсутствии генов вирулентности (строки) и названия штаммов Streptococcus (столбцы). Наконец, PathoProT передаст и обработает этот вывод с помощью наших собственных сценариев R для иерархической кластеризации (алгоритм полной связи) и создания тепловой карты для визуализации. Штаммы Streptococcus будут отсортированы на основе их профилей генов вирулентности (), и будет построено филогенетическое дерево, пользователи смогут оценить взаимосвязи между близкородственными S . mitis групповых видов / штаммов, а также соответствующие им гены вирулентности образуют заметные кластеры на дендрограммах. Таким образом, этот сравнительный анализ патогеномики может дать отличное представление о профилях генов вирулентности у разных видов Streptococcus . Например, не существует инструмента биоинформатики, который выполняет те же функции, что и PathoProT, а именно прогнозирует и позволяет сравнивать гены вирулентности в геномах разных видов бактерий.
Блок-схема PathoProT.
PathoProT в основном реализован с использованием сценариев Perl и R. Входными данными PathoProT будут списки генов для выбранных штаммов / геномов, и конвейер будет генерировать тепловую карту в конце процесса.
Чтобы продемонстрировать особенности или функциональные возможности PathoProT, мы представляем сравнительное исследование патогеномики среди S . mitis группируют бактерии, используя порог в 50% как для идентичности последовательностей, так и для покрытия, чтобы получить представление об их профилях генов вирулентности.На основании созданной тепловой карты PathoProT ряд предполагаемых генов вирулентности, по-видимому, сохраняется среди всех видов группы мититов (). Консервативные гены hasC (hasC1 или SMU . 322c) , которые кодируют UTP-глюкозо-1-фосфат уридилилтрансферазу (или UDP-пирофосфорилазу глюкозы) (M6Spy1871), участвуют в синтезе гиалуроновой кислоты вдоль (HA) капсулы. с двумя соседними генами: hasA и hasB в пределах имеет оперон.[21]. Фактически, это Streptococcus pneumoniae , наиболее патогенный вид S . Группа mitis обладает полисахаридной капсулой, которая способствует бактериальному патогенезу [22]. В Streptococcus HA обнаружен, поскольку материал капсулы стрептококка у некоторых видов является важным фактором вирулентности, эффективно маскируя бактерии от распознавания иммунной системой хозяина [23,24], а также защищая их от реактивных оксидов, выделяемых лейкоцитами [25] ].Кроме того, возможно, что ГК играет важную роль в прикреплении стрептококков группы мититов и колонизации эпителиальных клеток, что приводит к устойчивости бактерий к фагоцитозу макрофагами [26–28].
Информативная тепловая карта, созданная инструментом PathoProT.
(A) Список консервативных генов вирулентности, переносимых всеми видами группы mitis, и (B) гены, связанные с синтезом RGP, которые могут дифференцировать M Clade и S Clade. Наличие гена вирулентности было отмечено красным цветом, а отсутствие генов вирулентности было отмечено черным.
Другой консервативный ген вирулентности, slrA , кодирует стрептококковую липопротеин ротамазу А, которая является одним из основных поверхностных белков, экспрессируемых S . pneumoniae . Этот ген является важным циклофилином, который модулирует биологическую функцию белков вирулентности на первой стадии пневмококковой инфекции [29]. Вероятно, что ген slrA способствует инвазии клеток-хозяев и облегчает колонизацию пневмококков и присоединение к ним в S . бактерий группы mitis [30,31]. Кроме того, сообщалось, что дефицит slrA снижает вирулентность бактерий из-за его влияния на адгезию и интернализацию эпителиальными и эндотелиальными клетками [29]. Аналогичным образом, консервативный ген lmb кодирует ламинин-связывающий белок, который впервые был идентифицирован в Streptococcus agalactiae [32]. Практически идентичные адгезины позже были обнаружены как у Streptococcus suis [33], так и у Streptococcus pyogenes [34,35].Было предложено, что адгезины lmb помогают в бактериальном патогенезе посредством инвазии поврежденного эпителия [36]. В целом, многие поверхностные липопротеины и адгезины, которые важны для вирулентности и патогенных инфекций, в S очень консервативны. mitis группа бактерий.
Согласно филогенетическому дереву, сгенерированному в левой части тепловой карты PathoProT (), группу мититов можно четко разделить на две категории: S Clade ( S . sanguinis , S . гордонии , S . parasanguinis , S . Австралийский , S . cristatus и S . oligofermentans ) и M Clade ( S . mitis , S . infantis , S . tigurinus , S . oralis и S ). Это филогенетическое родство S . mitis групповых видов указывает на близкое родство между видами в M Clade и между видами в S Clade. Интересно, что мы обнаружили, что гены rgp могут использоваться для дифференциации двух разных кладов на тепловой карте. Например, эти маркерные гены присутствуют у всех видов S Clade, но отсутствуют у всех видов M Clade.
Кластер генов rgp (B, C, D, F и G) отвечает за синтез полисахарида рамнозы-глюкозы (RGP) в Streptococcus mutans .Примечательно, что подобные гены участвуют в синтезе рамнана у Escherichia coli [37]. Фактически, было предложено, чтобы E . coli и S . mutans имеют общий путь синтеза рамнана, основанный на их сходстве в синтезе RGP [37]. Функция rgpB заключается в переносе второго остатка рамнозы на остаток рамнозы на N -ацетилглюкозамине, связанном с липидным носителем, за которым следует rgpF , который позже катализирует перенос третьего остатка рамнозы на вторую рамнозу. остаток полученного гликолипидного носителя.И rgpB , и rgpF предположительно должны работать поочередно при удлинении цепи рамнана. Гомологичные рамнозилтрансферазы rgpB и rgpF были обнаружены у Streptococcus thermophilus (STER1436) и Streptococcus gordonii (SGO1022). С другой стороны, гены rgpC и rgpD кодируют предполагаемые переносчики ABC, специфичные для RGP (гомологичный STER1434 в S . thermophilus и гомологичный SGO1024 в S . gordonii ), которые играют роль в экспорте полисахаридов [37]. Ген rgpG (гомолог S . gordonii SGO1723) инициирует синтез RGP путем переноса N -ацетилглюкозамин-1-фосфата на липидный носитель [38].
Гены rgp также вовлечены в патогенез нескольких видов Streptococcus . Например, rgp играет важную роль в бактериальной вирулентности, а также вызывает воспалительную реакцию в S . suis [39]. Индукция инфекционного эндокардита по S . Сообщалось, что mutans запускается генами rgp посредством высвобождения оксида азота [40], агрегации тромбоцитов [41] и придания устойчивости к фагоцитозу полиморфно-ядерными лейкоцитами человека [42]. Следовательно, S Clade S . Виды группы mitis , которые продуцируют эти богатые рамнозой полимеры, могут демонстрировать патогенез, отличный от видов M Clade Streptococcus , с целью установления большей вирулентности и увеличения выживаемости в клетках-хозяевах.Недавнее исследование выявило группу стрептококков Sanguinis в качестве распространенного возбудителя транзиторной бактериемии, которая потенциально может привести к инфекционному эндокардиту. Сообщалось также, что эта группа присутствовала в нескольких случаях вирулентной сепсисной инфекции у пациентов с нейтропенией [43].
Инструменты поиска последовательности
Мы включили в StreptoBase два типа механизмов BLAST, стандартный BLAST и VFDB BLAST, для поиска штаммов Streptococcus , ближайших к штамму запроса. Эти эксклюзивные поиски BLAST функционально основаны на автономном инструменте BLAST [44], загруженном из NCBI. Оба механизма BLAST поддерживают три типа функций BLAST, а именно BLASTN, BLASTP и BLASTX. Пользователи могут определять полноту генома (%) и идентичность генома (%) в формах отправки инструментов BLAST. Эти специализированные инструменты BLAST призваны облегчить пользователям поиск сходства их запрашиваемых последовательностей с последовательностями генома Streptococcus , последовательностями генов (стандартный BLAST), а также с генами вирулентности VFDB (VFDB BLAST), что позволяет пользователям проверить, действительно ли их представляющие интерес гены являются генами потенциальной вирулентности с использованием подхода гомологии последовательностей.
Дальнейшая работа и заключение
С развитием технологии NGS будет секвенировано еще видов или штаммов Streptococcus , что создает острую потребность в хранении, просмотре, извлечении и анализе огромных объемов данных генома и разработке специализированных инструментов для сравнительный анализ этих геномов.
Здесь мы успешно описали и продемонстрировали функциональные возможности StreptoBase, в частности разработанные нами биоинформатические конвейеры для анализа геномных данных Streptococcus .
Эта специализированная биологическая база данных будет постоянно обновляться, чтобы предоставлять последние обновления генома и исследования, связанные с родом Streptococcus , а также обеспечивать точность и полезность S . mitis данные генома вида группы и аннотация. Мы ожидаем, что StreptoBase будет служить полезным ресурсом и платформой для анализа, особенно для сравнительного анализа S . mitis групповых геномов для исследовательских сообществ.Мы призываем других исследователей или исследовательские группы вносить предложения и делиться с нами своими аннотациями, мнениями и собранными данными по адресу ym.ude.mu@grig.
Группа геномных ресурсов и анализа Oral Streptococcus mitis
Функции базы данных и встроенные инструменты биоинформатики
S . mitis Виды группы являются важными колонизаторами ротовой полости и иногда связаны с серьезными инфекциями [15]. Кроме того, недавно было высказано предположение, что эти организмы играют важную роль в патогенезе гриппа [8].Таким образом, геномное исследование разнообразных S . mitis Бактерии группы необходимы для понимания того, как эти микроорганизмы переходят от комменсального образа жизни во рту к последующему патогенезу. Однако для широкого спектра S не существует специализированной базы данных генома. mitis групповых геномов для сравнительной геномики. Хотя большинство баз данных по биологическим геномам сосредоточены только на содержании генома и генетической изменчивости, мы определили необходимость создания функциональных биоинформатических инструментов для исследования детерминант вирулентности в геномах посредством сравнительной патогеномики, а также для сравнения содержания генома и генетической изменчивости в пределах S . mitis группа бактерий.
Инструмент парного сравнения генома (PGC)
Мы разработали и настроили веб-инструмент PGC для S . mitis группирует бактерии, что позволяет пользователям выбирать и проводить попарные сравнения между двумя выбранными пользователем геномами Streptococcus . Список геномов Streptococcus доступен в инструменте PGC StreptoBase, что позволяет пользователям выбрать два генома Streptococcus для сравнения штаммов или видов.Кроме того, пользователи могут загрузить свои собственные последовательности генома, нуклеотидов или белков, и сравнить их с геномами Streptococcus в StreptoBase.
Вкратце, конвейер PGC поддерживается NUCmer, который разработан для выравнивания полногеномных последовательностей, и Circos, который является хорошо зарекомендовавшим себя инструментом для визуализации генома. После того, как пользователи отправят свои задания на наш сервер, PGC вызовет программу NUCmer для согласования выбранных пользователем геномов, а внутренние сценарии будут использоваться для обработки выходных данных выравнивания генома и создания входных файлов, анализируемых в Circos, чтобы сгенерировать макет круговой идеограммы выравнивания.В отличие от обычного линейного отображения выравниваний, круговая компоновка показывает взаимосвязь между парами позиций с кариотипами и связями, кодирующими положение, размер и ориентацию связанных геномных элементов.
В веб-интерфейсе PGC предусмотрены три определяемых пользователем параметра, включая минимальный процент идентичности (%), порог слияния (bp) и порог связи (bp). Граница минимального процента идентичности определяет гомологичную область (представленную связями / лентами на графике Circos) между двумя сравниваемыми геномами.Порог слияния позволяет объединить две ссылки / ленты, расстояние между которыми находится в пределах определенного пользователем порога, а порог связи позволяет пользователям исключать любые сопоставленные / гомологичные области, размер генома которых меньше заданного пользователем порогового значения. Дорожка гистограммы добавляется во внешнем кольце кругового графика, чтобы указать процент картированных областей, что позволяет пользователям быстро идентифицировать потенциальные отступы (обозначенные белыми промежутками) и области картирования (обозначенные зелеными диаграммами) между двумя выровненными геномами.Реализация конвейера PGC управляется с помощью сценариев Perl. Этот конвейер производит два типа выходных данных: результаты выравнивания NUCmer и высококачественный график Circos (формат SVG). Пользователи могут бесплатно загрузить эти результаты для публикации или дальнейшего анализа на странице результатов PGC.
Существующий инструмент сравнения генома микробов (MGC) использует метод вычитания генома in silico для идентификации генетических элементов, специфичных для группы штаммов [16]. В то время как инструмент PGC использует файлы генома и NUCmer для выполнения попарного выравнивания генома, инструмент MGC использует in silico фрагментированные последовательности генома и выполняет BLASTN по группам запросов.Напротив, браузер VISTA, хорошо известный своим биологическим применением, может выполнять предварительно вычисленное попарное и множественное выравнивание генома с использованием как глобального, так и локального выравнивания [17]. В отличие от круговых графиков и гистограмм, которые генерируются инструментом PGC, результаты выравнивания, сгенерированные VISTA Browser, отображаются с использованием трека VISTA в формате графика, чтобы показать сохраненные области. Кроме того, основанный на Java инструмент сравнения Artemis (ACT) с открытым исходным кодом требует, чтобы пользователи сгенерировали файл сравнения, который идентифицирует области гомологии между сборочным и эталонным геномом с помощью таких программ, как BLASTN, TBLASTX или Mummer, которые должны быть загружены в ACT [18].Сравнительная визуализация ACT выполняется с использованием компонентов Artemis. Напротив, наш инструмент PGC обеспечивает однопоточный процесс попарного выравнивания генома и мгновенное отображение графика Circos сравнительного выравнивания.
Чтобы продемонстрировать полезность PGC, мы сравнили S . mitis B6 (полный геном) и 17/34 (черновой вариант генома) в качестве примера в.
Парное сравнение генома между S . mitis B6 и S . mitis 17/34 с использованием инструмента PGC, встроенного в StreptoBase.
50% идентичности последовательностей и 50% покрытия последовательностей использовали для сравнения штаммов с использованием инструмента PGC. A и B выделяют отступы парного сравнения генома между S . mitis B6 и S . mitis 17/34.
Параметры были установлены как 80% от минимального процента, значение по умолчанию — порог связи 1000 бит и порог слияния 2000 бит. S . mitis B6 был выделен в Германии, а S . mitis 17/34 был выделен из уретры российского пациента с уретритом.На основе сгенерированного графика PGC оба значения S . mitis геномов обычно имели высокое сходство, так как большинство их геномных областей могли быть выровнены (). Одной из особенностей графика PGC является его способность быстро определять предполагаемые отступы посредством визуализации пробелов в графике, что подтверждается информацией, отображаемой на дорожке гистограммы. Например, два разрыва () указывают на отсутствие геномных областей в S . mitis 17/34 геном.Внешняя круглая полоса графика показывает измерения размера генома, которые составляют примерно 2 МБ для обоих S . mitis геномов. Исходя из разрыва, наблюдаемого в (indel «A»), потеря гена произошла около позиции 400 000 п.н.
Затем мы исследовали гены, расположенные в индексе «A» в S . mitis B6 () путем визуализации этой области с помощью SGB. Мы идентифицировали множество генов, связанных с фагами, связанных с этой областью. Для дальнейшего изучения этой области мы использовали PHAST (инструмент поиска PHAge) для аннотирования и идентификации последовательностей профагов, обнаруженных в пределах S . mitis B6 геном (You Zhou et al., 2011). Интактный профаг размером 56 КБ с 82 CDS и содержанием GC 39,9% был обнаружен с 390 924 до 446 969 пар оснований. Начиная с S . mitis B6 — это полный геном, поэтому мы можем указать положение пары оснований непосредственно в нашем файле аннотации B6. Согласно результатам PHAST, это интактный профаг S . mitis B6 содержал фаг-ассоциированные гены, включая белок фаговой интегразы, фаговый CI-подобный репрессор, фаг-связывающий белок, портальный белок фага, белок морфогенеза головки фага семейства SPP1 и белки фагового капсида.Поэтому мы предлагаем S . mitis B6, возможно, недавно приобрел этот интактный профаг. Графическое изображение интактного профага с различными типами фаг-связанных генов показано на.
В S обнаружен интактный профаг. митис B6. Этот профаг имеет 85 предсказанных генов.
На основании инделя «B», обнаруженного на графике PGC, мы выявили неполный профаг размером 24 Кбайт с содержанием GC 39,17%, расположенный в положениях от 1356040 пар оснований до 1380128 пар оснований. Интересно, что эта область содержит полный оперон атр , регулируемый Белок CcpA в этом неполном профаге S . mitis геном B6. Гены оперона atp показаны в. Эти гены, кодирующие АТФ-синтазы, обычно используются оральными стрептококками для адаптации к кислой среде хозяина за счет создания более щелочной внутренней системы.
Таблица 5
Синтазы АТФ в опероне atp из S . митис B6.
Метка локуса
Имя гена
Функциональная аннотация
smi_1315
atpE
АТФ-синтаза C-цепочка (EC 3.6.3.14)
smi_1314
atpB
Цепь АТФ-синтазы А (EC 3.6.3.14)
smi_1313
atpF-синтаза Цепь B 901 3.14)
smi_1312
atpH
Дельта-цепь АТФ-синтазы (EC 3.6.3.14)
smi_1311
в цепи синтаза альфа-6.3.14)
smi_1310
atpG
Гамма-цепь АТФ-синтазы (EC 3.6.3.14)
smi_1309
beta. 3.14.Следовательно, возможно, что приобретение этого оперона atp осуществляется неполным профагом S . mitis B6 посредством горизонтального переноса генов помог его комменсальному статусу поддерживать оптимальный уровень pH для биоэнергетических процессов S . mitis клеток B6.
PathoProT был разработан для прогнозирования генов вирулентности путем сравнения аминокислотных последовательностей Streptococcus с базой данных факторов вирулентности (VFDB) [20].PathoProT использует автономные инструменты BLAST, загруженные с веб-сайта NCBI. VFDB (версия 2012) в настоящее время содержит набор из 19775 экспериментально подтвержденных генов вирулентности, происходящих от широкого круга различных видов бактерий, что обеспечивает полезный ресурс для поиска гомологии последовательностей. Пользователи могут выбрать список из штаммов Streptococcus для сравнительного анализа и установить пороговое значение, например, идентичность генома и полноту для поиска BLAST через нашу предоставленную онлайн-форму.Параметры по умолчанию для конвейера PathoProT установлены на 50% идентичности последовательностей и 50% полноты последовательности для поиска и идентификации генов ортологичной вирулентности в выбранных геномах Streptococcus . Однако пользователи могут применять желаемые отсечки для поиска гомологии, чтобы достичь оптимальных уровней строгости в своих анализах.
Вкратце, конвейер PathoProT в основном реализован с использованием Perl. Собственные сценарии Perl будут обрабатывать выходные данные BLAST (сгенерированные путем поиска этих последовательностей запросов в VFDB) для каждого предсказанного RAST белка (последовательности запроса) в выбранных пользователем геномах и определять предполагаемую вирулентность на основе заданных пользователем параметров.Отфильтрованные результаты BLAST объединяются и организуются в матричную таблицу, содержащую информацию о наличии или отсутствии генов вирулентности (строки) и названия штаммов Streptococcus (столбцы). Наконец, PathoProT передаст и обработает этот вывод с помощью наших собственных сценариев R для иерархической кластеризации (алгоритм полной связи) и создания тепловой карты для визуализации. Штаммы Streptococcus будут отсортированы на основе их профилей генов вирулентности (), и будет построено филогенетическое дерево, пользователи смогут оценить взаимосвязи между близкородственными S . mitis групповых видов / штаммов, а также соответствующие им гены вирулентности образуют заметные кластеры на дендрограммах. Таким образом, этот сравнительный анализ патогеномики может дать отличное представление о профилях генов вирулентности у разных видов Streptococcus . Например, не существует инструмента биоинформатики, который выполняет те же функции, что и PathoProT, а именно прогнозирует и позволяет сравнивать гены вирулентности в геномах разных видов бактерий.
Блок-схема PathoProT.
PathoProT в основном реализован с использованием сценариев Perl и R. Входными данными PathoProT будут списки генов для выбранных штаммов / геномов, и конвейер будет генерировать тепловую карту в конце процесса.
Чтобы продемонстрировать особенности или функциональные возможности PathoProT, мы представляем сравнительное исследование патогеномики среди S . mitis группируют бактерии, используя порог в 50% как для идентичности последовательностей, так и для покрытия, чтобы получить представление об их профилях генов вирулентности.На основании созданной тепловой карты PathoProT ряд предполагаемых генов вирулентности, по-видимому, сохраняется среди всех видов группы мититов (). Консервативные гены hasC (hasC1 или SMU . 322c) , которые кодируют UTP-глюкозо-1-фосфат уридилилтрансферазу (или UDP-пирофосфорилазу глюкозы) (M6Spy1871), участвуют в синтезе гиалуроновой кислоты вдоль (HA) капсулы. с двумя соседними генами: hasA и hasB в пределах имеет оперон.[21]. Фактически, это Streptococcus pneumoniae , наиболее патогенный вид S . Группа mitis обладает полисахаридной капсулой, которая способствует бактериальному патогенезу [22]. В Streptococcus HA обнаружен, поскольку материал капсулы стрептококка у некоторых видов является важным фактором вирулентности, эффективно маскируя бактерии от распознавания иммунной системой хозяина [23,24], а также защищая их от реактивных оксидов, выделяемых лейкоцитами [25] ].Кроме того, возможно, что ГК играет важную роль в прикреплении стрептококков группы мититов и колонизации эпителиальных клеток, что приводит к устойчивости бактерий к фагоцитозу макрофагами [26–28].
Информативная тепловая карта, созданная инструментом PathoProT.
(A) Список консервативных генов вирулентности, переносимых всеми видами группы mitis, и (B) гены, связанные с синтезом RGP, которые могут дифференцировать M Clade и S Clade. Наличие гена вирулентности было отмечено красным цветом, а отсутствие генов вирулентности было отмечено черным.
Другой консервативный ген вирулентности, slrA , кодирует стрептококковую липопротеин ротамазу А, которая является одним из основных поверхностных белков, экспрессируемых S . pneumoniae . Этот ген является важным циклофилином, который модулирует биологическую функцию белков вирулентности на первой стадии пневмококковой инфекции [29]. Вероятно, что ген slrA способствует инвазии клеток-хозяев и облегчает колонизацию пневмококков и присоединение к ним в S . бактерий группы mitis [30,31]. Кроме того, сообщалось, что дефицит slrA снижает вирулентность бактерий из-за его влияния на адгезию и интернализацию эпителиальными и эндотелиальными клетками [29]. Аналогичным образом, консервативный ген lmb кодирует ламинин-связывающий белок, который впервые был идентифицирован в Streptococcus agalactiae [32]. Практически идентичные адгезины позже были обнаружены как у Streptococcus suis [33], так и у Streptococcus pyogenes [34,35].Было предложено, что адгезины lmb помогают в бактериальном патогенезе посредством инвазии поврежденного эпителия [36]. В целом, многие поверхностные липопротеины и адгезины, которые важны для вирулентности и патогенных инфекций, в S очень консервативны. mitis группа бактерий.
Согласно филогенетическому дереву, сгенерированному в левой части тепловой карты PathoProT (), группу мититов можно четко разделить на две категории: S Clade ( S . sanguinis , S . гордонии , S . parasanguinis , S . Австралийский , S . cristatus и S . oligofermentans ) и M Clade ( S . mitis , S . infantis , S . tigurinus , S . oralis и S ). Это филогенетическое родство S . mitis групповых видов указывает на близкое родство между видами в M Clade и между видами в S Clade. Интересно, что мы обнаружили, что гены rgp могут использоваться для дифференциации двух разных кладов на тепловой карте. Например, эти маркерные гены присутствуют у всех видов S Clade, но отсутствуют у всех видов M Clade.
Кластер генов rgp (B, C, D, F и G) отвечает за синтез полисахарида рамнозы-глюкозы (RGP) в Streptococcus mutans .Примечательно, что подобные гены участвуют в синтезе рамнана у Escherichia coli [37]. Фактически, было предложено, чтобы E . coli и S . mutans имеют общий путь синтеза рамнана, основанный на их сходстве в синтезе RGP [37]. Функция rgpB заключается в переносе второго остатка рамнозы на остаток рамнозы на N -ацетилглюкозамине, связанном с липидным носителем, за которым следует rgpF , который позже катализирует перенос третьего остатка рамнозы на вторую рамнозу. остаток полученного гликолипидного носителя.И rgpB , и rgpF предположительно должны работать поочередно при удлинении цепи рамнана. Гомологичные рамнозилтрансферазы rgpB и rgpF были обнаружены у Streptococcus thermophilus (STER1436) и Streptococcus gordonii (SGO1022). С другой стороны, гены rgpC и rgpD кодируют предполагаемые переносчики ABC, специфичные для RGP (гомологичный STER1434 в S . thermophilus и гомологичный SGO1024 в S . gordonii ), которые играют роль в экспорте полисахаридов [37]. Ген rgpG (гомолог S . gordonii SGO1723) инициирует синтез RGP путем переноса N -ацетилглюкозамин-1-фосфата на липидный носитель [38].
Гены rgp также вовлечены в патогенез нескольких видов Streptococcus . Например, rgp играет важную роль в бактериальной вирулентности, а также вызывает воспалительную реакцию в S . suis [39]. Индукция инфекционного эндокардита по S . Сообщалось, что mutans запускается генами rgp посредством высвобождения оксида азота [40], агрегации тромбоцитов [41] и придания устойчивости к фагоцитозу полиморфно-ядерными лейкоцитами человека [42]. Следовательно, S Clade S . Виды группы mitis , которые продуцируют эти богатые рамнозой полимеры, могут демонстрировать патогенез, отличный от видов M Clade Streptococcus , с целью установления большей вирулентности и увеличения выживаемости в клетках-хозяевах.Недавнее исследование выявило группу стрептококков Sanguinis в качестве распространенного возбудителя транзиторной бактериемии, которая потенциально может привести к инфекционному эндокардиту. Сообщалось также, что эта группа присутствовала в нескольких случаях вирулентной сепсисной инфекции у пациентов с нейтропенией [43].
Инструменты поиска последовательности
Мы включили в StreptoBase два типа механизмов BLAST, стандартный BLAST и VFDB BLAST, для поиска штаммов Streptococcus , ближайших к штамму запроса.Эти эксклюзивные поиски BLAST функционально основаны на автономном инструменте BLAST [44], загруженном из NCBI. Оба механизма BLAST поддерживают три типа функций BLAST, а именно BLASTN, BLASTP и BLASTX. Пользователи могут определять полноту генома (%) и идентичность генома (%) в формах отправки инструментов BLAST. Эти специализированные инструменты BLAST призваны облегчить пользователям поиск сходства их запрашиваемых последовательностей с последовательностями генома Streptococcus , последовательностями генов (стандартный BLAST), а также с генами вирулентности VFDB (VFDB BLAST), что позволяет пользователям проверить, действительно ли их представляющие интерес гены являются генами потенциальной вирулентности с использованием подхода гомологии последовательностей.
Дальнейшая работа и заключение
С развитием технологии NGS будет секвенировано еще видов или штаммов Streptococcus , что создает острую потребность в хранении, просмотре, извлечении и анализе огромных объемов данных генома и разработке специализированных инструментов для сравнительный анализ этих геномов.
Здесь мы успешно описали и продемонстрировали функциональные возможности StreptoBase, в частности разработанные нами биоинформатические конвейеры для анализа геномных данных Streptococcus .
Эта специализированная биологическая база данных будет постоянно обновляться, чтобы предоставлять последние обновления генома и исследования, связанные с родом Streptococcus , а также обеспечивать точность и полезность S . mitis данные генома вида группы и аннотация. Мы ожидаем, что StreptoBase будет служить полезным ресурсом и платформой для анализа, особенно для сравнительного анализа S . mitis групповых геномов для исследовательских сообществ.Мы призываем других исследователей или исследовательские группы вносить предложения и делиться с нами своими аннотациями, мнениями и собранными данными по адресу ym.ude.mu@grig.
Streptococcus oralis — обзор
Дисбиоз
Многие антибиотики также оказывают негативное влияние на комменсальную микробиоту. Очень восприимчивые бактерии, такие как оральный Neisseria spp. подавляются широким спектром противомикробных агентов, в то время как оральные стрептококки viridans группы mitis , такие как Streptococcus mitis и Streptococcus oralis, 38 и другие необычные оральные комменсальные микробиоты, такие как Rothia mucilaginosa Capnocytophaga spp., вероятно, будут выбраны противомикробными агентами, к которым бактерии лишь незначительно восприимчивы, если вообще чувствительны. В частности, пенициллины, рифампицин, клиндамицин, макролиды, бацитрацин и ванкомицин значительно снижают устойчивость к колонизации, вероятно, потому, что они ингибируют грамположительные неспорулирующие бактерии, продуцирующие молочную кислоту, такие как бифидобактерии. 39 Некоторые цефалоспорины также вредны, тогда как триметоприм-сульфаметоксазол и хинолоны были объявлены «дружественными», поэтому их часто используют в качестве профилактики. 39 Неожиданно использование имипенема в более высоких дозах привело к увеличению случаев диареи, вызванной Clostridium difficile. 40 Новые антибиотики, такие как тигециклин, обладающие активностью против анаэробных грамположительных бактерий, могут привести к увеличению числа пациентов с Candida. Это изменение обычно отражает заметные нарушения экологии кишечника. 41 Однако этот эффект не увеличивает риск заражения C.difficile. 42 Хлоргексидиновые жидкости для полоскания рта, используемые для уменьшения образования зубного налета и гингивита, также влияют на микрофлору. 43-45
Поскольку нормальная комменсальная микробиота прикрепляется к поверхностям эпителия, их потеря создает экологический вакуум, который позволяет другим организмам устанавливать колонизацию, занимая свободные поверхности клеток или используя избыток питательных веществ. Коллапс экологии неизменно проявляется в чрезмерном росте дрожжей и колонизации нозокомиальными бактериями, такими как Klebsiella pneumoniae и P.aeruginosa 46-49 и невозможность прямого или косвенного обнаружения жизнеспособных анаэробов. 47,48 Примеры инфекционных осложнений, связанных с нарушением нормального микробного равновесия, включают выделение ранее необычных видов, таких как Enterococcus faecium и Clostridium septicum.
Диспепсия является достаточно обычным явлением для регулярного назначения антацидов, таких как блокаторы гистамина-2 (h3) и ингибиторы протонной помпы.Пониженная кислотность желудочного сока непреднамеренно разрушает естественный барьер, который предотвращает колонизацию желудка и кишечника оральной комменсальной микробиотой, многие из которых устойчивы к большинству антимикробных агентов, используемых для профилактики у больных с нарушениями здоровья. Когда пациенты проглатывают большое количество слизи в результате тяжелого мукозита, любая оральная комменсальная микробиота может пережить попадание в кишечник. Таким образом, потеря желудочного барьера эффективно расширяет область потенциальных мест для колонизации на всю длину пищеварительного тракта, что может объяснить патогенез α-гемолитической (группа viridans) стрептококковой бактериемии. 50,51,52 Стрептококки Viridans, обычно S. mitis, , могут вызывать инфекции, связанные с опасными для жизни осложнениями, включая септический шок, пневмонит и острый респираторный дистресс-синдром. Высокие дозы цитарабина предрасполагают к этим осложнениям. 51,53-55 Наконец, экология кишечной микробиоты заметно изменяется из-за повреждения кишечника, вызванного лечением некоторыми цитостатическими агентами, такими как цитарабин, 56,57 , реакцией трансплантат против хозяина, 58 и облучение всего тела. 59 Различные коагулазонегативные стафилококки, включая Staphylococcus epidermidis, , также присутствуют в эндогенной микробиоте полости рта и желудочно-кишечном тракте пациентов с нейтропенией. 60 Анализ плазмидного спектра изолятов коагулазонегативных стафилококков кровотока показал, что слизистая оболочка является источником бактериемии у 70% пациентов, находящихся в гематологическом отделении одного центра. 61 Другие обнаружили, что сосудистые катетеры были наиболее вероятным источником коагулазонегативной бактериемии Staphylococcus у пациентов с гематологическими злокачественными новообразованиями (см. Главу 302).Однако есть четкие доказательства того, что коагулазонегативные стафилококки, ответственные за бактериемию, могут происходить как из слизистой оболочки ротовой полости и кишечника, так и из катетера. 62,63
Идентификация Streptococcus pneumoniae и других стрептококков Mitis: важность молекулярных методов
1.
Facklam R (2002) Что случилось со стрептококками: обзор таксономических и номенклатурных изменений. Clin Microbiol Rev 15: 613–630
PubMed
PubMed Central
Google ученый
2.
Jensen A, Scholz CFP, Kilian M (2016) Переоценка таксономии группы мититов рода Streptococcus на основе филогенетического анализа всего генома и предложенная реклассификация Streptococcus dentisani как Streptococcus subis . dentisani греб. nov., Streptococcus tigurinus как Streptococcus oralis subsp. tigurinus греб. nov. и Streptococcus oligofermentans как более поздний синоним Streptococcus cristatus .Int J Syst Evol Microbiol 66: 4803–4820. https://doi.org/10.1099/ijsem.0.001433
CAS
Статья
PubMed
Google ученый
3.
Patel S, Gupta RS (2018) Четкое разграничение четырнадцати различных групп видов в пределах рода Streptococcus на основе филогении на основе генома и молекулярных сигнатур. Заразить Genet Evol 66: 130–151. https://doi.org/10.1016/j.meegid.2018.09.020
CAS
Статья
PubMed
Google ученый
4.
Troeger C, Forouzanfar M, Rao PC, Khalil I, Brown A, Swartz S, Fullman N, Mosser J, Thompson RL, Reiner RC Jr, Abajobir A, Alam N, Alemayohu MA, Amare AT, Antonio CA, Asayesh H, Авокпахо Э, Барак А., Бешир М.А., Бонея Д.Д., Брауэр М., Дандона Л., Дандона Р., Фитчетт Дж.РА, Гебрехивот Т.Т., Хайлу, Великобритания, Хотез П.Дж., Касаеян А., Ходжа Т., Киссун Н., Книббс Л., Кумар Г.А., Рай Р.К. , El Razek HMA, Mohammed MSK, Nielson K, Oren E, Osman A, Patton G, Qorbani M, Roba HS, Sartorius B, Savic M, Shigematsu M, Sykes B, Swaminathan S, Topor-Madry R, Ukwaja K, Werdecker A, Yonemoto N, El Sayed ZM, Lim SS, Naghavi M, Vos T., Hay SI, Murray CJL, Mokdad AH (2017) Оценки глобальной, региональной и национальной заболеваемости, смертности и этиологии инфекций нижних дыхательных путей в 195 стран: систематический анализ глобального бремени болезней, исследование 2015 г.Lancet Infect Dis 17: 1133–1161. https://doi.org/10.1016/S1473-3099(17)30396-1
Статья
Google ученый
5.
Валь Б., О’Брайен К.Л., Гринбаум А., Маджумдер А., Лю Л., Чу И., Лукшич И., Наир Х., Макаллистер Д.А., Кэмпбелл Х., Рудан И., Блэк Р., Нолл, доктор медицины (2018) Бёрден of Streptococcus pneumoniae и Haemophilus influenzae Болезнь типа b у детей в эпоху конъюгированных вакцин: глобальные, региональные и национальные оценки за 2000-15 годы.Lancet Glob Health 6: e744 – e757
PubMed
PubMed Central
Google ученый
6.
van Aalst M, Lötsch F, Spijker R, van der Meer JTM, Langendam MW, Goorhuis A, Grobusch MP, de Bree GJ (2018) Заболеваемость инвазивным пневмококковым заболеванием у пациентов с ослабленным иммунитетом: систематический обзор и мета -анализ. Travel Med Infect Dis 24: 89–100. https://doi.org/10.1016/j.tmaid.2018.05.016
Статья
PubMed
Google ученый
7.
Cherazard R, Epstein M, Doan TL, Salim T, Bharti S, Smith MA (2017) Устойчивость к противомикробным препаратам Streptococcus pneumoniae : распространенность, механизмы и клинические последствия. Am J Ther 24: e361 – e369. https://doi.org/10.1097/MJT.0000000000000551
Статья
PubMed
Google ученый
8.
Weinberger DM, Malley R, Lipsitch M (2011) Замена серотипа при заболевании после пневмококковой вакцинации: обсуждение доказательств.Ланцет 378: 1962–1973. https://doi.org/10.1016/S0140-6736(10)62225-8
Статья
PubMed
PubMed Central
Google ученый
9.
Koelman DLH, Brouwer MC, van de Beek D (2020) Возникновение пневмококкового менингита в Европе и Северной Америке. Clin Microbiol Infect 26: 199–204. https://doi.org/10.1016/j.cmi.2019.04.032
CAS
Статья
PubMed
Google ученый
10.
Doern CD, Burnham CAD (2010) Непросто быть зеленым: стрептококки группы viridans с акцентом на педиатрические клинические проявления. J Clin Microbiol 48: 3829–3835. https://doi.org/10.1128/JCM.01563-10
Мердок Д.Р., Кори Г.Р., Хоэн Б., Миро Дж. М., Фаулер В. Г. мл., Байер А. С., Карчмер А. В., Олейсон Л., Паппас П. А., Морейллон П., Чемберс СТ, Чу В. Х., Фалько В., Холланд Д. Д., Джонс П., Кляйн Д. Л., Raymond NJ, Read KM, Tripodi MF, Utili R, Wang A, Woods CW, Cabell CH, International Collaboration on Endocarditis-Prospective Cohort Study (ICE-PCS) Investigators (2009) Клиническая презентация, этиология и исход инфекционного эндокардита в 21 st век: международное сотрудничество по проспективному когортному исследованию эндокардита.Arch Intern Med 169: 463–473. https://doi.org/10.1001/archinternmed.2008.603
Статья
PubMed
PubMed Central
Google ученый
13.
Shelburne SA, Sahasrabhojane P, Saldana M, Yao H, Su X, Horstmann N, Thompson E, Flores AR (2014) Streptococcus mitis штаммы, вызывающие тяжелое клиническое заболевание у онкологических больных. Emerg Infect Dis 20: 762–771. https://doi.org/10.3201/eid2005.130953
Статья
PubMed
PubMed Central
Google ученый
14.
Mundy LS, Janoff EN, Schwebke KE, Shanholtzer CJ, Willard KE (1998) Неопределенность в идентификации Streptococcus pneumoniae. Оптохин, растворимость в желчи, подавление и ДНК-зонд AccuProbe. Am J Clin Pathol 109: 55–61
CAS
PubMed
Google ученый
15.
Martín-Galiano AJ, Balsalobre L, Fenoll A, de la Campa AG (2003) Молекулярная характеристика ассоциированных с заболеванием стрептококков из группы митита, которые чувствительны к оптохинам.Противомикробные агенты Chemother 47: 3187–3194
PubMed
PubMed Central
Google ученый
16.
Arbique JC, Poyart C, Trieu-Cuot P, Quesne G, da Gloria CM, Steigerwalt AG, Morey RE, Jackson D, Davidson RJ, Facklam RR (2004) Точность фенотипического и генотипического тестирования для идентификации Streptococcus pneumoniae и описание Streptococcus pseudopneumoniae sp. ноя J Clin Microbiol 42: 4686–4696
CAS
PubMed
PubMed Central
Google ученый
17.
Balsalobre L, Hernández-Madrid A, Llull D, Martín-Galiano AJ, García E, Fenoll A, de la Campa AG (2006) Молекулярная характеристика ассоциированных с заболеванием стрептококков из группы мититов, чувствительных к оптохинам. J Clin Microbiol 44: 4163–4171
CAS
PubMed
PubMed Central
Google ученый
18.
Richter SS, Heilmann KP, Dohrn CL, Riahi F, Beekmann SE, Doern GV (2008) Точность фенотипических методов идентификации изолятов Streptococcus pneumoniae , включенных в программы надзора.J Clin Microbiol 46: 2184–2188. https://doi.org/10.1128/JCM.00461-08
Das A, Behera B, Madan M, Mathur P, Pandey A (2010) Эмпиема, вызванная чувствительной к оптохину Streptococcus mitis в ходе ветряной оспы. Индийский J Pediatr 77: 464. https://doi.org/10.1007/s12098-010-0024-2
Статья
PubMed
Google ученый
21.
Simões AS, Sá-Leão R, Eleveld MJ, Tavares DA, Carriço JA, Bootsma HJ, Hermans PW (2010) Высокоустойчивые к пенициллину полирезистентные пневмококкоподобные штаммы, колонизирующие детей в Оейрасе, Португалия: геномный характеристики и значение для наблюдения.J Clin Microbiol 48: 238–246. https://doi.org/10.1128/JCM.01313-09
Rolo D, Simões SA, Domenech A, Fenoll A, Liñares J, de Lencastre H, Ardanuy C, Sá-Leão R (2013) Изоляты болезней Streptococcus pseudopneumoniae и нетипичных S. pneumoniae , предположительно идентифицированных как атипичные S. pneumoniae в Испании. PLoS One 8: e57047. https://doi.org/10.1371/journal.pone.0057047
CAS
Статья
PubMed
PubMed Central
Google ученый
24.
Simões AS, Tavares DA, Rolo D, Ardanuy C, Goossens H, Henriques-Normark B, Linares J, de Lencastre H, Sá-Leão R (2016) lytA методы идентификации могут неверно идентифицировать Streptococcus pneumoniae .Диагностика Microbiol Infect Dis 85: 141–148. https://doi.org/10.1016/j.diagmicrobio.2016.03.018
CAS
Статья
PubMed
Google ученый
25.
Yahiaoui RY, den Heijer CD, Wolfs P, Bruggeman CA, Stobberingh EE (2016) Оценка фенотипических и молекулярных методов идентификации Streptococcus pneumoniae . Future Microbiol 11: 43–50. https://doi.org/10.2217/fmb.15.124
CAS
Статья
PubMed
Google ученый
26.
Sadowy E, Bojarska A, Kuch A, Skoczyńska A, Jolley KA, Maiden MCJ, van Tonder AJ, Hammerschmidt S, Hryniewicz W. (2020) Отношения между стрептококками из группы мититов, ошибочно идентифицированных как Streptococcus pneumoniae . Eur J Clin Microbiol Infect Dis. https://doi.org/10.1007/s10096-020-03916-6 [онлайн перед печатью]
Donati C, Hiller NL, Tettelin H, Muzzi A, Croucher NJ, Angiuoli SV, Oggioni M, Dunning Hotopp JC, Hu FZ, Riley DR, Covacci A, Mitchell TJ, Bentley SD, Kilian M, Ehrlich GD, Rappuoli R, Moxon ER, Masignani V (2010) Структура и динамика пангенома Streptococcus pneumoniae и близкородственных видов. Геном Биол 11: R107.dx.doi.org. https://doi.org/10.1186/gb-2010-11-10-r107
CAS
Статья
PubMed
PubMed Central
Google ученый
29.
Sanguinetti L, Toti S, Reguzzi V, Bagnoli F, Donati C (2012) Новый вычислительный метод идентифицирует события внутри- и межвидовой рекомбинации у Staphylococcus aureus и Streptococcus pneumoniae . PLoS Comput Biol 8: e1002668 dx.doi.org. https://doi.org/10.1371/journal.pcbi.1002668
CAS
Статья
PubMed
PubMed Central
Google ученый
30.
Шерман Дж. М. (1937) Стрептококки. Бактериол Ред. 1: 3–97
CAS
PubMed
PubMed Central
Google ученый
31.
Bentley RW, Leigh JA, Collins MD (1991) Внутриродовая структура Streptococcus на основе сравнительного анализа последовательностей малых субъединиц рРНК.Int J Syst Bacteriol 41: 487–494
CAS
PubMed
Google ученый
32.
Kawamura Y, Hou XG, Sultana F, Miura H, Ezaki T (1995) Определение последовательностей 16S рРНК Streptococcus mitis и Streptococcus gordonii cus и филогенетических связей между членами рода Int J Syst Bacteriol 45: 406–408
CAS
PubMed
Google ученый
33.
Hardie JM, Whiley RA (1997) Классификация и обзор Streptococcus и Enterococcus . J Appl Microbiol Symp Sup 83: 1S – 11S
CAS
Google ученый
34.
Ruoff KL, Whiley RA, Beighton D (2003) Streptococcus. В: Murray PR, Baron EJ, Jorgensen JH, Pfaller MA, Yolken RH (eds) Руководство по клинической микробиологии, 8-е изд. ASM Press, Вашингтон, округ Колумбия, стр. 405–421
Google ученый
35.
Lund E, Henrichsen J (1978) Глава XI Лабораторная диагностика, серология и эпидемиология Streptococcus pneumoniae . Методы Microbiol 12: 241–262
Google ученый
36.
Филлипс Дж., Баркер Р., Броган О. (1988) Оптохин-резистентный Streptococcus pneumoniae . Ланцет 2: 281
CAS
PubMed
Google ученый
37.
Pinto TC, Souza AR, de Pina SE, Costa NS, Borges Neto AA, Neves FP, Merquior VL, Dias CA, Peralta JM, Teixeira LM (2013) Фенотипическая и молекулярная характеристика оптохин-устойчивого стрептококка pneumoniae из Бразилии, с описанием пяти новых мутаций в гене atpC .J Clin Microbiol 51: 3242–3249. https://doi.org/10.1128/JCM.01168-13
CAS
Статья
PubMed
PubMed Central
Google ученый
38.
Сацке К., Тернер П., Виролайнен-Юлкунен А., Адриан П. В., Антонио М., Харе К. М., Энао-Рестрепо А. М., Лич А. Дж., Клугман К. П., Портер Б. Д., Са-Леан Р., Скотт Дж. А., Нохинек Х. , О’Брайен К.Л., Рабочая группа ВОЗ по пневмококковому носительству (2013 г.) Стандартный метод определения носительства в верхних дыхательных путях Streptococcus pneumoniae : обновленные рекомендации Рабочей группы Всемирной организации здравоохранения по пневмококковому носительству.Vaccine 32: 165–179. https://doi.org/10.1016/j.vaccine.2013.08.062
Статья
PubMed
Google ученый
39.
Slotved HC, Facklam RR, Fuursted K (2017) Оценка нового теста растворимости желчи и MALDI-TOF для дифференциации Streptococcus pneumoniae от других стрептококков группы мититов. Sci Rep 7: 7167. https://doi.org/10.1038/s41598-017-07772-x
CAS
Статья
PubMed
PubMed Central
Google ученый
40.
Teles C, Smith A, Ramage G, Lang S (2011) Идентификация клинически значимых стрептококков группы viridans с помощью фенотипического и генотипического анализа. Eur J Clin Microbiol Infect Dis 30: 243–250. https://doi.org/10.1007/s10096-010-1076-y
CAS
Статья
PubMed
Google ученый
41.
Имаи К., Немото Р., Кодана М., Тарумото Н., Сакаи Дж., Кавамура Т., Икебути К., Мицутаке К., Мураками Т., Маэсаки С., Фудзивара Т., Хаякава С., Хосино Т., Секи М., Маеда Т. (2020) Быстрая и точная идентификация стрептококков группы мититов с помощью секвенатора MinION Nanopore.Front Cell Infect Microbiol 10:11. https://doi.org/10.3389/fcimb.2020.00011
CAS
Статья
PubMed
PubMed Central
Google ученый
42.
Murray PR (2010) Матричная лазерная десорбционная ионизационная времяпролетная масс-спектрометрия: полезность для таксономии и эпидемиологии. Clin Microbiol Infect 16: 1626–1630. https://doi.org/10.1111/j.1469-0691.2010.03364.x
CAS
Статья
PubMed
Google ученый
43.
Икрянникова Л.Н., Филимонова А.В., Малахова М.В., Савинова Т., Филимонова О., Ильина Е.Н., Дубовицкая В.А., Сидоренко С.В., Говорун В.М. (2012) Дискриминация между Streptococcus pneumoniae и Streptococcus mitis по масс-спектру на основе сортировки по масс-спектру Mitis . Clin Microbiol Infect 19: 1066–1071. https://doi.org/10.1111/1469-0691.12113
CAS
Статья
Google ученый
44.
Zhou M, Yang Q, Kudinha T, Zhang L, Xiao M, Kong F, Zhao Y, Xu YC (2016) Использование матричной лазерной десорбционной ионизации-время пролета (MALDI-TOF) с дополнением секвенирование генов 16S рРНК и gyrB для практической идентификации клинически важных стрептококков группы Viridans (VGS).Front Microbiol 7: 1328. https://doi.org/10.3389/fmicb.2016.01328
Статья
PubMed
PubMed Central
Google ученый
45.
Harju I, Lange C, Kostrzewa M, Maier T, Rantakokko-Jalava K, Haanperä M (2017) Улучшенная дифференциация Streptococcus pneumoniae и других S. mitis группы стрептококков, улучшенных с помощью MALDI Bi Содержимое базы данных MALDI Biotyper и новый алгоритм интерпретации результатов.J Clin Microbiol 55: 914–922. https://doi.org/10.1128/JCM.01990-16
Skov Sørensen UB, Yao K, Yang Y, Tettelin H, Kilian M (2016) Экспрессия капсульного полисахарида в комменсале Streptococcus видов: генетическое и антигенное сходство с Streptococcus pneumoniae .mBio 7: e01844 – e01816. https://doi.org/10.1128/mBio.01844-16
Статья
PubMed
PubMed Central
Google ученый
48.
Lessa FC, Milucky J, Rouphael NG, Bennett NM, Talbot HK, Harrison LH, Farley MM, Walston J, Pimenta F, Gertz RE, Rajam G, da Gloria CM, Beall B, Whitney CG (2018 ) Streptococcus mitis , экспрессирующий капсулу пневмококка серотипа 1. Sci Rep 8: 17959. https://doi.org/10.1038/s41598-018-35921-3
CAS
Статья
PubMed
PubMed Central
Google ученый
49.
Пимента Ф, Герц Р. Младший, Пак Ш., Ким Э, Моура I, Милаки Дж., Руфаэль Н., Фарли М. М., Харрисон Л. Х., Беннетт Н. М., Бигого Дж., Фейкин Д. Р., Брейман Р., Лесса ФК, Уитни К. Г., Раджам Дж., Schiffer J, da Gloria CM, Beall B (2019) Streptococcus infantis, Streptococcus mitis и Streptococcus oralis с очень похожими локусами cps5 и антигенным родством с пневмококками серотипа 5. Front Microbiol 9: 3199. https://doi.org/10.3389/fmicb.2018.03199
Статья
PubMed
PubMed Central
Google ученый
50.
Slotved HC, Kaltoft M, Skovsted IC, Kerrn MB, Espersen F (2004) Простой, быстрый тест латексной агглютинации для серотипирования пневмококков (Pneumotest-Latex). J Clin Microbiol 42: 2518–2225
CAS
PubMed
PubMed Central
Google ученый
51.
Kuch A, Gołębiewska A, Waśko I, Ronkiewicz P, Markowska M, Hryniewicz W., Skoczyńska A (2014) Полезность Pneumotest-latex для прямого серотипирования образцов Streptococcus pneumoniae .J Clin Microbiol 52: 2647–2649. https://doi.org/10.1128/JCM.00451-14
Hilty M, Wüthrich D, Salter SJ, Engel H, Campbell S, Sá-Leão R, de Lencastre H, Hermans P, Sadowy E, Turner P, Chewapreecha C, Diggle M, Pluschke G, McGee L , Eser ÖK, Low DE, Smith-Vaughan H, Endimiani A, Küffer M, Dupasquier M, Beaudoing E, Weber J, Bruggmann R, Hanage WP, Parkhill J, Hathaway LJ, Mühlemann K, Bentley SD (2014) Глобальный филогеномный анализ из неинкапсулированных Streptococcus pneumoniae обнаруживает классическую линию с глубоким ветвлением, которая отличается от множества спорадических линий.Genome Biol Evol 6: 3281–3294. https://doi.org/10.1093/gbe/evu263
CAS
Статья
PubMed
PubMed Central
Google ученый
54.
Varghese R, Jayaraman R, Veeraraghavan B (2017) Текущие проблемы точной идентификации Streptococcus pneumoniae и его серогрупп / серотипов в эпоху вакцин. J Microbiol Methods 141: 48–54. https://doi.org/10.1016/j.mimet.2017.07.015
CAS
Статья
PubMed
Google ученый
55.
da Gloria CM, Tondella ML, McCaustland K, Weidlich L, McGee L, Mayer LW, Steigerwalt A, Whaley M, Facklam RR, Fields B, Carlone G, Ades EW, Dagan R, Sampson JS (2007) Оценка и улучшение ПЦР в реальном времени, нацеленная на гены lytA , ply и psaA для обнаружения пневмококковой ДНК. J Clin Microbiol 45: 2460–2466
Google ученый
56.
Book M, Lehmann LE, Zhang X, Stüber F (2013) Мониторинг инфекции: от посева крови до полимеразной цепной реакции (ПЦР).Лучшая практика Res Clin Anaesthesiol 27: 279–288. https://doi.org/10.1016/j.bpa.2013.06.010
Статья
PubMed
Google ученый
57.
da Gloria CM, Pimenta FC, Jackson D, Roundtree A, Ahmad Y, Millar EV, O’Brien KL, Whitney CG, Cohen AL, Beall BW (2010) Возвращение к переносу пневмококка с использованием обогащения бульона и Методы ПЦР для улучшенного выявления носительства и серотипов. J Clin Microbiol 48: 1611–1618. https: // doi.org / 10.1128 / JCM.02243-09
CAS
Статья
Google ученый
58.
Strålin K, Herrmann B, Abdeldaim G, Olcén P, Holmberg H, Mölling P (2014) Сравнение образцов мокроты и носоглоточного аспирата, а также мишеней гена ПЦР lytA и Spn9802 для быстрой количественной ПЦР пневмококковой пневмонии. J Clin Microbiol 52: 83–89. https://doi.org/10.1128/JCM.01742-13
CAS
Статья
PubMed
PubMed Central
Google ученый
59.
Gillis HD, Lang ALS, ElSherif M, Martin I, Hatchette TF, McNeil SA, LeBlanc JJ, Сеть по надзору за серьезными исходами (SOS) Канадского исследования иммунизации (CIRN) (2017) Оценка диагностической точности обнаружения вирусов с помощью ПЦР Streptococcus pneumoniae из мазков из носоглотки, собранных для вирусных исследований у взрослых канадцев, госпитализированных с внебольничной пневмонией: исследование сети по надзору за серьезными исходами (SOS) Канадского исследования иммунизации (CIRN). BMJ Open 7: e015008.https://doi.org/10.1136/bmjopen-2016-015008
Статья
PubMed
PubMed Central
Google ученый
60.
Sundell N, Andersson LM, Brittain-Long R, Sundvall PD, Alsiö Å, Lindh M, Gustavsson L, Westin J (2019) ПЦР-определение респираторных патогенов у бессимптомных и симптоматических взрослых. J Clin Microbiol 57: e00716 – e00718. https://doi.org/10.1128/JCM.00716-18
Статья
PubMed
PubMed Central
Google ученый
61.
Virolainen A, Salo P, Jero J, Karma P, Eskola J, Leinonen M (1994) Сравнение анализа ПЦР с бактериальной культурой для обнаружения Streptococcus pneumoniae в жидкости среднего уха у детей с острым средним отитом. J Clin Microbiol 32: 2667–2670
CAS
PubMed
PubMed Central
Google ученый
62.
Сало П., Ортквист А., Лейнонен М. (1995) Диагностика бактериемической пневмококковой пневмонии путем амплификации фрагмента гена пневмолизина в сыворотке.J Infect Dis 171: 479–482
CAS
PubMed
Google ученый
63.
Kaijalainen T, Rintamäki S, Herva E, Leinonen M (2002) Оценка генно-технологических и традиционных методов идентификации Streptococcus pneumoniae . J Microbiol Methods 51: 111–118
CAS
PubMed
Google ученый
64.
Whatmore AM, Efstratiou A, Pickerill AP, Broughton K, Woodard G, Sturgeon D, George R, Dowson CG (2000) Генетические отношения между клиническими изолятами Streptococcus pneumoniae, Streptococcus oralis, и Streptococcus m. : характеристика «атипичных» пневмококков и организмов, родственных S.mitis , несущий генов, кодирующих фактор вирулентности S. pneumoniae . Infect Immun 68: 1374–1382
CAS
PubMed
PubMed Central
Google ученый
65.
Neeleman C, Klaassen CH, Klomberg DM, de Valk HA, Mouton JW (2004) Пневмолизин является ключевым фактором в ошибочной идентификации макролид-устойчивого Streptococcus pneumoniae и является предполагаемым фактором вирулентности S. mitis. и другие стрептококки.J Clin Microbiol 42: 4355–4357
CAS
PubMed
PubMed Central
Google ученый
66.
Johnston C, Hinds J, Smith A, van der Linden M, Van Eldere J, Mitchell TJ (2010) Обнаружение большого количества генов пневмококковой вирулентности у стрептококков группы мититов. J Clin Microbiol 48: 2762–2769. https://doi.org/10.1128/JCM.01746-09
CAS
Статья
PubMed
PubMed Central
Google ученый
67.
Mosser JL, Tomasz A (1970) Холин-содержащая тейхоевая кислота как структурный компонент клеточной стенки пневмококка и ее роль в чувствительности к лизису автолитическим ферментом. J Biol Chem 245: 287–298
CAS
PubMed
Google ученый
68.
Gillespie SH, Ullman C, Smith MD, Emery V (1994) Обнаружение Streptococcus pneumoniae в образцах мокроты с помощью ПЦР. J Clin Microbiol 32: 1308–1311
CAS
PubMed
PubMed Central
Google ученый
69.
Llull D, López R, García E (2006) Характерные сигнатуры гена lytA обеспечивают основу для быстрой и надежной диагностики инфекций Streptococcus pneumoniae . J Clin Microbiol 44: 1250–1256
CAS
PubMed
PubMed Central
Google ученый
70.
Tavares DA, Handem S, Carvalho RJ, Paulo AC, de Lencastre H, Hinds J, Sá-Leão R (2019) Идентификация Streptococcus pneumoniae с помощью анализа ПЦР в реальном времени, нацеленного на SP2020.Научный журнал 9: 3285. https://doi.org/10.1038/s41598-019-39791-1
CAS
Статья
PubMed
PubMed Central
Google ученый
71.
Morrison KE, Lake D, Crook J, Carlone GM, Ades E, Facklam R, Sampson JS (2000) Подтверждение psaA во всех 90 серотипах Streptococcus pneumoniae с помощью ПЦР и потенциала этого анализа для идентификации и диагностики. J Clin Microbiol 38: 434–437
CAS
PubMed
PubMed Central
Google ученый
72.
Verhelst R, Kaijalainen T, De Baere T, Verschraegen G, Claeys G, Van Simaey L, De Ganck C, Vaneechoutte M (2003) Сравнение пяти генотипических методов идентификации оптохин-устойчивых пневмококкоподобных изолятов. J Clin Microbiol 41: 3521–3525
CAS
PubMed
PubMed Central
Google ученый
73.
Messmer TO, Sampson JS, Stinson A, Wong B, Carlone GM, Facklam RR (2004) Сравнение четырех анализов полимеразной цепной реакции на специфичность идентификации Streptococcus pneumoniae .Диагностика Microbiol Infect Dis 49: 249–254
CAS
PubMed
Google ученый
74.
Jado I, Fenoll A, Casal J, Pérez A (2001) Идентификация гена psaA , кодирующего пневмококковый поверхностный адгезин A, у стрептококков группы viridans, кроме Streptococcus pneumoniae . Clin Diagn Lab Immunol 8: 895–898
CAS
PubMed
PubMed Central
Google ученый
75.
Zhang Q, Ma Q, Su D, Li Q, Yao W, Wang C (2010) Идентификация горизонтального переноса гена и рекомбинации гена PsaA в группе Streptococcus mitis . Microbiol Immunol 54: 313–319. https://doi.org/10.1111/j.1348-0421.2010.00216.x
CAS
Статья
PubMed
Google ученый
76.
Park HK, Lee SJ, Yoon JW, Shin JW, Shin HS, Kook JK, Myung SC, Kim W (2010) Идентификация гена cpsA как специфического маркера для распознавания Streptococcus pneumoniae из стрептококков группы viridans.J Med Microbiol 59: 1146–1152. https://doi.org/10.1099/jmm.0.017798-0
da Gloria CM, Pimenta FC, Moura I, Roundtree A, Gertz RE Jr, Li Z, Jagero G, Bigogo G, Junghae M, Conklin L, Feikin DR, Breiman RF, Whitney CG, Beall BW (2013) Непневмококковый Стрептококки группы mitis затрудняют обнаружение пневмококковых капсульных серотип-специфических локусов в верхних дыхательных путях. PeerJ 1: e97. https://doi.org/10.7717/peerj.97
CAS
Статья
Google ученый
79.
Whalan RH, Funnell SG, Bowler LD, Hudson MJ, Robinson A, Dowson CG (2006) Распределение и генетическое разнообразие липопротеинов-переносчиков ABC PiuA и PiaA в пределах Streptococcus pneumoniae и родственных стрептококков.J Bacteriol 188: 1031–1038
CAS
PubMed
PubMed Central
Google ученый
80.
Wyllie AL, Pannekoek Y, Bovenkerk S, van Engelsdorp GJ, Ferwerda B, van de Beek D, Sanders EAM, Trzciński K, van der Ende A (2017). различать Streptococcus pneumoniae и другие виды стрептококков. Откройте Biol 7: 170074. https://doi.org/10.1098/rsob.170074
CAS
Статья
PubMed
PubMed Central
Google ученый
81.
Peela SCM, Sistla S, Tamilarasu K, Krishnamurthy S, Adhishivam B (2018) Проверка гена приобретения пневмококкового железа ( piaA ) для точной идентификации Streptococcus pneumoniae . Индийский журнал J Med Microbiol 36: 504–507. https://doi.org/10.4103/ijmm.IJMM_18_274
Статья
PubMed
Google ученый
82.
Zbinden A, Köhler N, Bloemberg GV (2011) ПЦР-анализ на основе recA для точной дифференциации Streptococcus pneumoniae от других стрептококков viridans.J Clin Microbiol 49: 523–527. https://doi.org/10.1128/JCM.01450-10
CAS
Статья
PubMed
PubMed Central
Google ученый
83.
Sistek V, Boissinot M, Boudreau DK, Huletsky A, Picard FJ, Bergeron MG (2012) Разработка ПЦР-анализа в реальном времени для специфического обнаружения и идентификации Streptococcus pseudopneumoniae с использованием recA ген. Clin Microbiol Infect 18: 1089–1096. https: // doi.org / 10.1111 / j.1469-0691.2011.03684.x
CAS
Статья
PubMed
Google ученый
84.
Scholz CF, Poulsen K, Kilian M (2012) Новый молекулярный метод идентификации Streptococcus pneumoniae , применимый к клинической микробиологии и исследованиям микробиома на основе последовательности 16S рРНК. J Clin Microbiol 50: 1968–1973. https://doi.org/10.1128/JCM.00365-12
CAS
Статья
PubMed
PubMed Central
Google ученый
85.
Suzuki N, Seki M, Nakano Y, Kiyoura Y, Maeno M, Yamashita Y (2005) Дискриминация Streptococcus pneumoniae от стрептококков группы viridans с помощью геномной субтрактивной гибридизации. J Clin Microbiol 43: 4528–4534
CAS
PubMed
PubMed Central
Google ученый
86.
Suzuki N, Yuyama M, Maeda S, Ogawa H, Mashiko K, Kiyoura Y (2006) Генотипическая идентификация предположительного Streptococcus pneumoniae с помощью ПЦР с использованием четырех генов, высокоспецифичных для S.пневмония . J Med Microbiol 55: 709–714
CAS
PubMed
Google ученый
87.
Wessels E, Schelfaut JJ, Bernards AT, Claas EC (2012) Оценка нескольких биохимических и молекулярных методов для идентификации Streptococcus pneumoniae и Streptococcus pseudopneumoniae и их обнаружения в респираторных образцах. J Clin Microbiol 50: 1171–1177. https://doi.org/10.1128/JCM.06609-11
CAS
Статья
PubMed
PubMed Central
Google ученый
88.
Croxen MA, Lee TD, Azana R, Hoang LM (2018) Использование геномики для разработки диагностического анализа для различения между Streptococcus pneumoniae и Streptococcus pseudopneumoniae . Microb Genom 4. https://doi.org/10.1099/mgen.0.000175
89.
Donati C, Hiller NL, Tettelin H, Muzzi A, Croucher NJ, Angiuoli SV, Oggioni M, Dunning Hotopp JC, Hu FZ , Riley DR, Covacci A, Mitchell TJ, Bentley SD, Kilian M, Ehrlich GD, Rappuoli R, Moxon ER, Masignani V (2010) Структура и динамика пангенома Streptococcus pneumoniae и близкородственных видов.Геном Биол 11: R107. https://doi.org/10.1186/gb-2010-11-10-r107
Вельско И.М., Перес М.С., Ричардс В.П. (2019) Выявление филогенетических взаимосвязей для Streptococcus mitis и Streptococcus oralis с помощью анализа ядра и пан-генома. Genome Biol Evol 11: 1077–1087. https://doi.org/10.1093/gbe/evz049
CAS
Статья
PubMed
PubMed Central
Google ученый
92.
Kilian M, Tettelin H (2019) Идентификация свойств, связанных с вирулентностью, путем сравнительного анализа генома Streptococcus pneumoniae, S.pseudopneumoniae, S. mitis , три подвида S. oralis и S. infantis . mBio 10: e01985 – e01919. https://doi.org/10.1128/mBio.01985-19
CAS
Статья
PubMed
PubMed Central
Google ученый
93.
Salvadori G, Junges R, Morrison DA, Petersen FC (2019) Компетенция в отношении Streptococcus pneumoniae и близких комменсальных родственников: механизмы и последствия. Front Cell Infect Microbiol 9:94.https://doi.org/10.3389/fcimb.2019.00094
CAS
Статья
PubMed
PubMed Central
Google ученый
94.
Gonzales-Siles L, Karlsson R, Schmidt P, Salvà-Serra F, Jaén-Luchoro D, Skovbjerg S, Moore ERB, Gomila M (2020) Подход пангенома для распознавания уникальных для видов маркеров генов для идентификации Streptococcus pneumoniae и Streptococcus pseudopneumoniae . Front Cell Infect Microbiol 10: 222.https://doi.org/10.3389/fcimb.2020.00222
Статья
PubMed
PubMed Central
Google ученый
95.
Poyart C, Quesne G, Coulon S, Berche P, Trieu-Cuot P (1998) Идентификация стрептококков до видового уровня путем секвенирования гена, кодирующего марганец-зависимую супероксиддисмутазу. J Clin Microbiol 36: 41–47
CAS
PubMed
PubMed Central
Google ученый
96.
Kawamura Y, Whiley RA, Shu SE, Ezaki T, Hardie JM (1999) Генетические подходы к идентификации группы мититов в пределах рода Streptococcus . Микробиология 145: 2605–2613
CAS
PubMed
Google ученый
97.
Drancourt M, Roux V, Fournier PE, Raoult D (2004) rpoB Идентификация на основе последовательности гена аэробных грамположительных кокков родов Streptococcus, Enterococcus, Gemella, Abiotrophia, и Granulica .J Clin Microbiol 42: 497–504
CAS
PubMed
PubMed Central
Google ученый
98.
Picard FJ, Ke D, Boudreau DK, Boissinot M, Huletsky A, Richard D, Ouellette M, Roy PH, Bergeron MG (2004) Использование последовательностей tuf для определения родоспецифической ПЦР и филогенетического анализа 28 видов стрептококков. J Clin Microbiol 42: 3686–3695
CAS
PubMed
PubMed Central
Google ученый
99.
Galloway-Peña J, Sahasrabhojane P, Tarrand J, Han XY, Shelburne SA (2014) Полиморфизмы gyrB точно определяют инвазивные виды стрептококков группы viridans. J Clin Microbiol 52: 2905–2912. https://doi.org/10.1128/JCM.01068-14
CAS
Статья
PubMed
PubMed Central
Google ученый
100.
Rasmunssen LH, Dargis R, Hojholt K, Christensen JJ, Skovgaard O, Justensen US, Rosenvinge FS, Moser C, Lukjancenko O, Rasmunssen S, Nielsen XC (2016) Секвенирование всего генома как инструмент филогенетического анализа клинических штаммов стрептококков группы мититов.Eur J Clin Microbiol Infect Dis 35: 1615–1625. https://doi.org/10.1007/s10096-016-2700-2
Статья
Google ученый
101.
Westling K, Julander I, Ljungman P, Vondracek M, Wretlind B, Jalal S (2008) Идентификация видов стрептококков группы viridans в изолятах клинической культуры крови путем анализа последовательности гена РНКазы P РНК, rnpB . J Inf Secur 56: 204–210
Google ученый
102.
Park HK, Yoon JW, Shin JW, Kim JY, Kim W (2010) rpoA — полезный ген для идентификации и классификации Streptococcus pneumoniae из близкородственных стрептококков группы viridans. FEMS Microbiol Lett 305: 58–64. https://doi.org/10.1111/j.1574-6968.2010.01913.x
CAS
Статья
PubMed
Google ученый
103.
Глазунова О.О., Рауль Д., Ру В. (2010) Частичное секвенирование гена recN : новый инструмент для идентификации и филогении в пределах рода Streptococcus .Int J Syst Evol Microbiol 60: 2140–2148. https://doi.org/10.1099/ijs.0.018176-0
CAS
Статья
PubMed
Google ученый
104.
Maeda Y, Goldsmith CE, Coulter WA, Mason C, Dooley JS, Lowery CJ, Millar BC, Moore JE (2011) Сравнение пяти локусов генов ( rnpB , 16S рРНК, 16S-23S рРНК, sodA и dnaJ ), чтобы помочь молекулярной идентификации стрептококков группы viridans и пневмококков.Br J Biomed Sci 68: 190–196
CAS
PubMed
Google ученый
105.
Enright MC, Spratt BG (1998) Схема мультилокусного типирования последовательностей для Streptococcus pneumoniae : идентификация клонов, связанных с серьезным инвазивным заболеванием. Микробиология 144: 3049–3060
CAS
PubMed
Google ученый
106.
Бишоп CJ, Aanensen DM, Jordan GE, Kilian M, Hanage WP, Spratt BG (2009) Отнесение штаммов к видам бактерий через Интернет.BMC Biol 7: 3. https://doi.org/10.1186/1741-7007-7-3
CAS
Статья
PubMed
PubMed Central
Google ученый
107.
Hanage WP, Kaijalainen T, Herva E, Saukkoriipi A, Syrjänen R, Spratt BG (2005) Использование данных мультилокусной последовательности для определения пневмококка. J Bacteriol 187: 6223–6230
Hanage WP, Fraser C, Spratt BG (2006) Последовательности, кластеры последовательностей и виды бактерий. Philos Trans R Soc Lond Ser B Biol Sci 361: 1917–1927. https://doi.org/10.1098/rstb.2006.1917
Статья
Google ученый
110.
Fuursted K, Littauer PJ, Greve T, Scholz CF (2016) Септицемия с Streptococcus pseudopneumoniae : отчет о трех случаях с очевидной ассоциацией печеночных или желчных протоков. Infect Dis (Лондон) 48: 636–639. https://doi.org/10.3109/23744235.2016.1157896
Статья
Google ученый
111.
Wood DE, Salzberg SL (2014) Kraken: сверхбыстрая классификация метагеномных последовательностей с использованием точного выравнивания. Геном Биол 15: R46.https://doi.org/10.1186/gb-2014-15-3-r46
Статья
PubMed
PubMed Central
Google ученый
112.
Zhang XX, Guo LY, Liu LL, Shen A, Feng WY, Huang WH, Hu HL, Hu B, Guo X, Chen TM, Chen HY, Jiang YQ, Liu G (2019) Диагностическое значение метагеномного секвенирования нового поколения для идентификации Streptococcus pneumoniae при бактериальном менингите у детей. BMC Infect Dis 19: 495. https://doi.org/10.1186/s12879-019-4132-y
Статья
PubMed
PubMed Central
Google ученый
113.
Jolley KA, Bliss CM, Bennett JS, Bratcher HB, Brehony C, Colles FM, Wimalarathna H, Harrison OB, Sheppard SK, Cody AJ, Maiden MC (2012) Рибосомное мультилокусное типирование последовательностей: универсальная характеристика бактерий от домена к штамму. Микробиология 158: 1005–1015. https://doi.org/10.1099/mic.0.055459-0


 Иногда может быть обнаружен на слизистой глотки и в фекалиях, куда попадают транзитом. При этом надо помнить, что этот микроб вносит существенный вклад в процессы, связанные с кариесом, занимая в последующем ведущее место в ассоциациях микроорганизмов, выделяемых из гнойного экссудата при воспалительных процессах в челюстно-лицевой области, включая одонтогенные абсцессы мозга и других органов. Кроме того, они же могут быть выделены их крови и сердечных клапанов больных инфекционным эндокардитом.
Иногда может быть обнаружен на слизистой глотки и в фекалиях, куда попадают транзитом. При этом надо помнить, что этот микроб вносит существенный вклад в процессы, связанные с кариесом, занимая в последующем ведущее место в ассоциациях микроорганизмов, выделяемых из гнойного экссудата при воспалительных процессах в челюстно-лицевой области, включая одонтогенные абсцессы мозга и других органов. Кроме того, они же могут быть выделены их крови и сердечных клапанов больных инфекционным эндокардитом. Иногда может быть обнаружен на слизистой глотки и в фекалиях, куда попадают транзитом. При этом надо помнить, что этот микроб вносит существенный вклад в процессы, связанные с кариесом, занимая в последующем ведущее место в ассоциациях микроорганизмов, выделяемых из гнойного экссудата при воспалительных процессах в челюстно-лицевой области, включая одонтогенные абсцессы мозга и других органов. Кроме того, они же могут быть выделены их крови и сердечных клапанов больных инфекционным эндокардитом.
Иногда может быть обнаружен на слизистой глотки и в фекалиях, куда попадают транзитом. При этом надо помнить, что этот микроб вносит существенный вклад в процессы, связанные с кариесом, занимая в последующем ведущее место в ассоциациях микроорганизмов, выделяемых из гнойного экссудата при воспалительных процессах в челюстно-лицевой области, включая одонтогенные абсцессы мозга и других органов. Кроме того, они же могут быть выделены их крови и сердечных клапанов больных инфекционным эндокардитом. Этот микроб является кариесогенным, а значит, может в дальнейшем вызывать бактериемию и бактериальный эндокардит у пациентов, не занимающихся регулярной санацией кариозных очагов в полости рта.
Этот микроб является кариесогенным, а значит, может в дальнейшем вызывать бактериемию и бактериальный эндокардит у пациентов, не занимающихся регулярной санацией кариозных очагов в полости рта.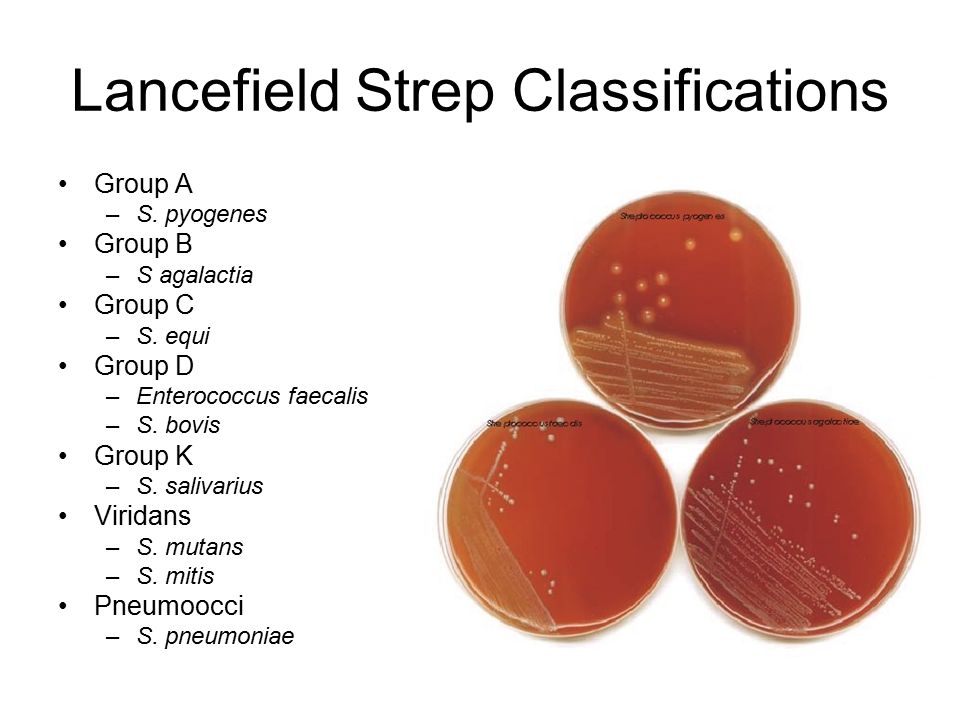
 Таким образом об этих группах мы даже не вспоминаем.
Таким образом об этих группах мы даже не вспоминаем.
 , Брико Н.И., Намазова-Баранова Л.С., Ряпис Л.А. Стрептококки и пневмококки. Ростов н/Д: Феникс, 2013.
, Брико Н.И., Намазова-Баранова Л.С., Ряпис Л.А. Стрептококки и пневмококки. Ростов н/Д: Феникс, 2013.
 J Clin Microbiol. 2017; 55(3): 914-22. doi: 10.1128/JCM.01990-16.
J Clin Microbiol. 2017; 55(3): 914-22. doi: 10.1128/JCM.01990-16. Species-specific difference in antimicrobial susceptibility among viridans group streptococci. Ann Lab Med. 2015; 35(2): 205-11. doi: 10.3343/alm.2015.35.2.205.
Species-specific difference in antimicrobial susceptibility among viridans group streptococci. Ann Lab Med. 2015; 35(2): 205-11. doi: 10.3343/alm.2015.35.2.205.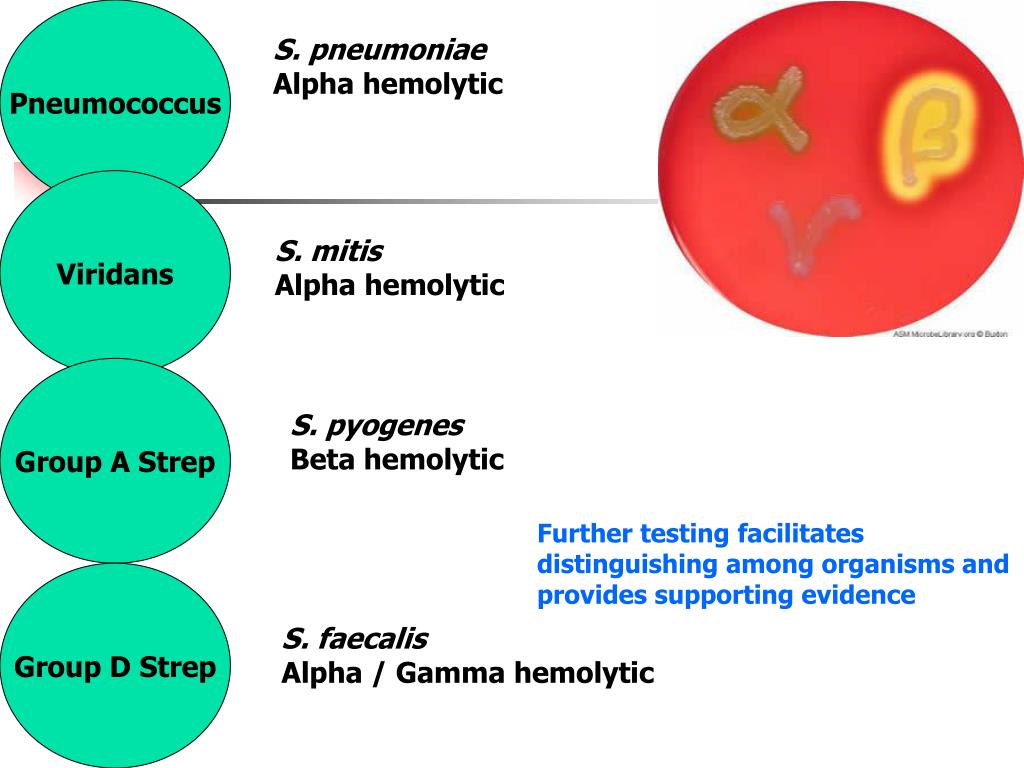 А., Алябьева Н.М., Пономаренко О.А., Куличенко Т.В., Артемова И.В., Лазарева А.В. и др. Динамика распространенности серотипов и антибиотикорезистентности носоглоточных пневмококков, выделенных у детей в 2010-2016 гг.: результаты ретроспективного когортного исследования. Вопросы современной педиатрии. 2017; 16 (5): 413-23. doi: 10.15690/vsp.v16i5.1806).
А., Алябьева Н.М., Пономаренко О.А., Куличенко Т.В., Артемова И.В., Лазарева А.В. и др. Динамика распространенности серотипов и антибиотикорезистентности носоглоточных пневмококков, выделенных у детей в 2010-2016 гг.: результаты ретроспективного когортного исследования. Вопросы современной педиатрии. 2017; 16 (5): 413-23. doi: 10.15690/vsp.v16i5.1806). Prevalence rate and antibiotic susceptibility of oral viridans group streptococci (VGS) in healthy children population. Adv Med Sci. 2006; 51 Suppl 1: 191-5.
Prevalence rate and antibiotic susceptibility of oral viridans group streptococci (VGS) in healthy children population. Adv Med Sci. 2006; 51 Suppl 1: 191-5. Crossing the barrier: evolution and spread of a major class of mosaic pbp2x in S. pneumoniae, S. mitis and S oralis. Int J Med Microbiol. 2007; 297: 503-12. https://doi.org/10.1016/j.ijmm.
Crossing the barrier: evolution and spread of a major class of mosaic pbp2x in S. pneumoniae, S. mitis and S oralis. Int J Med Microbiol. 2007; 297: 503-12. https://doi.org/10.1016/j.ijmm. Antimicrob Agents Chemother. 2017; 61(5). pii: e02646-16. doi: 10.1128/AAC.02646-16.
Antimicrob Agents Chemother. 2017; 61(5). pii: e02646-16. doi: 10.1128/AAC.02646-16. Мы будем очень благодарны, если Вы порекомендуете нас своим друзьям в социальных сетях.
Мы будем очень благодарны, если Вы порекомендуете нас своим друзьям в социальных сетях.
 agalactiae)»
agalactiae)» Наиболее высок уровень колонизации у женщин репродуктивного возраста и беременных женщин. У беременных СГВ выделяют из перечисленных областей в 7-30% случаев (чаще у молодых, имеющих высокую половую активность или пользующихся внутриматочными средствами контрацепции). Из них 60-75% остаются СГВ-носителями до конца беременности.
Наиболее высок уровень колонизации у женщин репродуктивного возраста и беременных женщин. У беременных СГВ выделяют из перечисленных областей в 7-30% случаев (чаще у молодых, имеющих высокую половую активность или пользующихся внутриматочными средствами контрацепции). Из них 60-75% остаются СГВ-носителями до конца беременности. agalactiae)
agalactiae)  [b/UZZeJR+ђαǃ;2+R$IӔO’ϥn`pvf’HZF/_w8;|sߺG?ON~w»?9;?w4 .oN/L{!~r:(ڽU;
qT
M0’&A:;poNw?_ȃSzR{n0 ߎ P:i:_RݔA9A[|ہ4SP»2VaH]CSj@ӕs%P/b:(o`hJ2ݽ{Y~+e
[b/UZZeJR+ђαǃ;2+R$IӔO’ϥn`pvf’HZF/_w8;|sߺG?ON~w»?9;?w4 .oN/L{!~r:(ڽU;
qT
M0’&A:;poNw?_ȃSzR{n0 ߎ P:i:_RݔA9A[|ہ4SP»2VaH]CSj@ӕs%P/b:(o`hJ2ݽ{Y~+e mitis группа бактерий.
mitis группа бактерий. В отличие от обычного линейного отображения выравниваний, круговая компоновка показывает взаимосвязь между парами позиций с кариотипами и связями, кодирующими положение, размер и ориентацию связанных геномных элементов.
В отличие от обычного линейного отображения выравниваний, круговая компоновка показывает взаимосвязь между парами позиций с кариотипами и связями, кодирующими положение, размер и ориентацию связанных геномных элементов.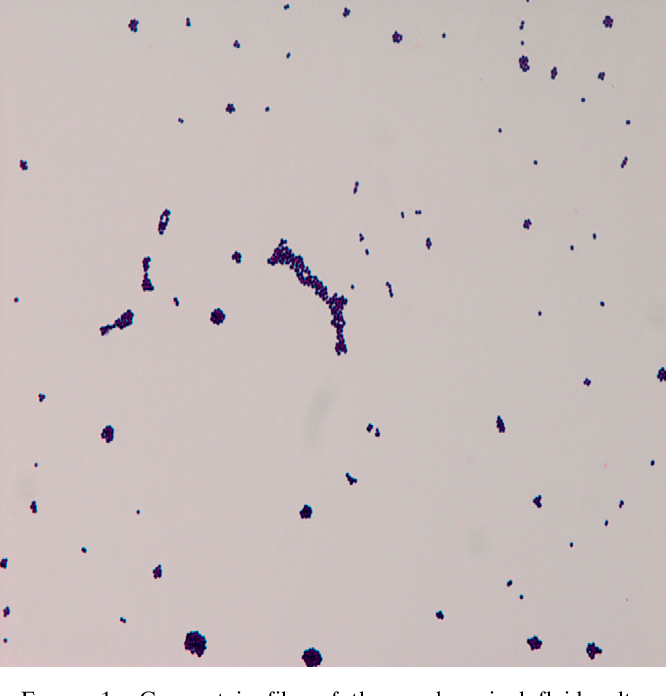 Реализация конвейера PGC управляется с помощью сценариев Perl. Этот конвейер производит два типа выходных данных: результаты выравнивания NUCmer и высококачественный график Circos (формат SVG). Пользователи могут бесплатно загрузить эти результаты для публикации или дальнейшего анализа на странице результатов PGC.
Реализация конвейера PGC управляется с помощью сценариев Perl. Этот конвейер производит два типа выходных данных: результаты выравнивания NUCmer и высококачественный график Circos (формат SVG). Пользователи могут бесплатно загрузить эти результаты для публикации или дальнейшего анализа на странице результатов PGC. Кроме того, основанный на Java инструмент сравнения Artemis (ACT) с открытым исходным кодом требует, чтобы пользователи сгенерировали файл сравнения, который идентифицирует области гомологии между сборочным и эталонным геномом с помощью таких программ, как BLASTN, TBLASTX или Mummer, которые должны быть загружены в ACT [18].Сравнительная визуализация ACT выполняется с использованием компонентов Artemis. Напротив, наш инструмент PGC обеспечивает однопоточный процесс попарного выравнивания генома и мгновенное отображение графика Circos сравнительного выравнивания.
Кроме того, основанный на Java инструмент сравнения Artemis (ACT) с открытым исходным кодом требует, чтобы пользователи сгенерировали файл сравнения, который идентифицирует области гомологии между сборочным и эталонным геномом с помощью таких программ, как BLASTN, TBLASTX или Mummer, которые должны быть загружены в ACT [18].Сравнительная визуализация ACT выполняется с использованием компонентов Artemis. Напротив, наш инструмент PGC обеспечивает однопоточный процесс попарного выравнивания генома и мгновенное отображение графика Circos сравнительного выравнивания. A и B выделяют отступы парного сравнения генома между S . mitis B6 и S . mitis 17/34.
A и B выделяют отступы парного сравнения генома между S . mitis B6 и S . mitis 17/34.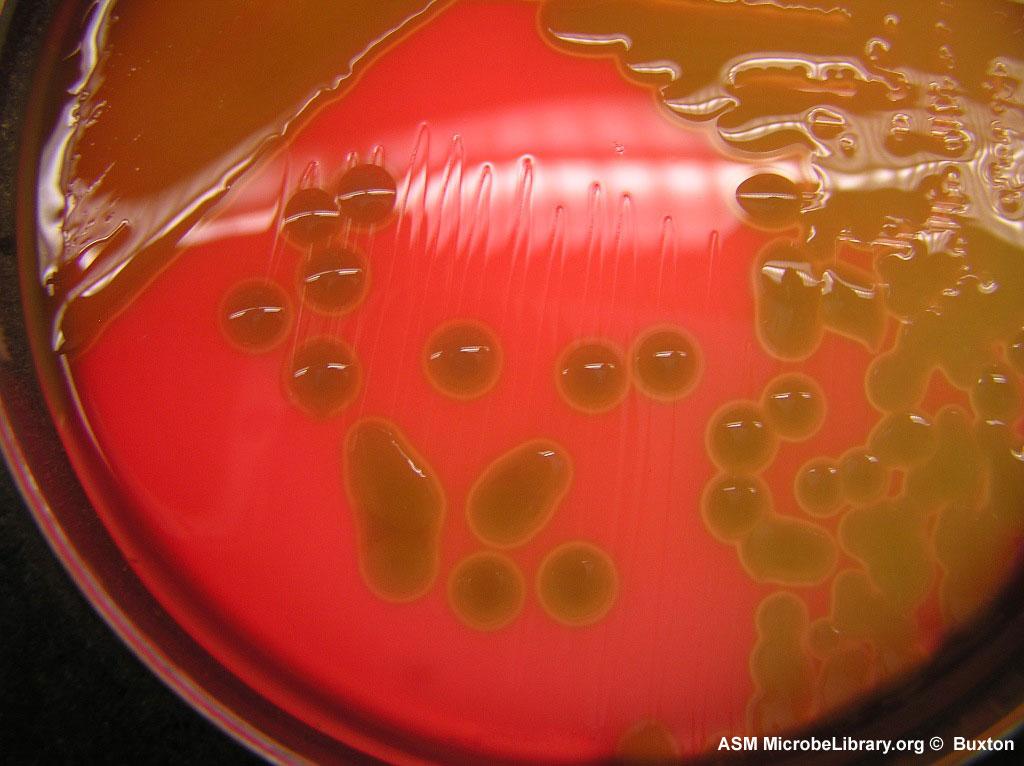 mitis геномов. Исходя из разрыва, наблюдаемого в (indel «A»), потеря гена произошла около позиции 400 000 п.н.
mitis геномов. Исходя из разрыва, наблюдаемого в (indel «A»), потеря гена произошла около позиции 400 000 п.н. Поэтому мы предлагаем S . mitis B6, возможно, недавно приобрел этот интактный профаг. Графическое изображение интактного профага с различными типами фаг-связанных генов показано на.
Поэтому мы предлагаем S . mitis B6, возможно, недавно приобрел этот интактный профаг. Графическое изображение интактного профага с различными типами фаг-связанных генов показано на. митис B6.
митис B6.  6.3.14)
6.3.14) PathoProT использует автономные инструменты BLAST, загруженные с веб-сайта NCBI. VFDB (версия 2012) в настоящее время содержит набор из 19775 экспериментально подтвержденных генов вирулентности, происходящих от широкого круга различных видов бактерий, что обеспечивает полезный ресурс для поиска гомологии последовательностей. Пользователи могут выбрать список из штаммов Streptococcus для сравнительного анализа и установить пороговое значение, например, идентичность генома и полноту для поиска BLAST через нашу предоставленную онлайн-форму.Параметры по умолчанию для конвейера PathoProT установлены на 50% идентичности последовательностей и 50% полноты последовательности для поиска и идентификации генов ортологичной вирулентности в выбранных геномах Streptococcus . Однако пользователи могут применять желаемые отсечки для поиска гомологии, чтобы достичь оптимальных уровней строгости в своих анализах.
PathoProT использует автономные инструменты BLAST, загруженные с веб-сайта NCBI. VFDB (версия 2012) в настоящее время содержит набор из 19775 экспериментально подтвержденных генов вирулентности, происходящих от широкого круга различных видов бактерий, что обеспечивает полезный ресурс для поиска гомологии последовательностей. Пользователи могут выбрать список из штаммов Streptococcus для сравнительного анализа и установить пороговое значение, например, идентичность генома и полноту для поиска BLAST через нашу предоставленную онлайн-форму.Параметры по умолчанию для конвейера PathoProT установлены на 50% идентичности последовательностей и 50% полноты последовательности для поиска и идентификации генов ортологичной вирулентности в выбранных геномах Streptococcus . Однако пользователи могут применять желаемые отсечки для поиска гомологии, чтобы достичь оптимальных уровней строгости в своих анализах. Собственные сценарии Perl будут обрабатывать выходные данные BLAST (сгенерированные путем поиска этих последовательностей запросов в VFDB) для каждого предсказанного RAST белка (последовательности запроса) в выбранных пользователем геномах и определять предполагаемую вирулентность на основе заданных пользователем параметров.Отфильтрованные результаты BLAST объединяются и организуются в матричную таблицу, содержащую информацию о наличии или отсутствии генов вирулентности (строки) и названия штаммов Streptococcus (столбцы). Наконец, PathoProT передаст и обработает этот вывод с помощью наших собственных сценариев R для иерархической кластеризации (алгоритм полной связи) и создания тепловой карты для визуализации. Штаммы Streptococcus будут отсортированы на основе их профилей генов вирулентности (), и будет построено филогенетическое дерево, пользователи смогут оценить взаимосвязи между близкородственными S . mitis групповых видов / штаммов, а также соответствующие им гены вирулентности образуют заметные кластеры на дендрограммах.
Собственные сценарии Perl будут обрабатывать выходные данные BLAST (сгенерированные путем поиска этих последовательностей запросов в VFDB) для каждого предсказанного RAST белка (последовательности запроса) в выбранных пользователем геномах и определять предполагаемую вирулентность на основе заданных пользователем параметров.Отфильтрованные результаты BLAST объединяются и организуются в матричную таблицу, содержащую информацию о наличии или отсутствии генов вирулентности (строки) и названия штаммов Streptococcus (столбцы). Наконец, PathoProT передаст и обработает этот вывод с помощью наших собственных сценариев R для иерархической кластеризации (алгоритм полной связи) и создания тепловой карты для визуализации. Штаммы Streptococcus будут отсортированы на основе их профилей генов вирулентности (), и будет построено филогенетическое дерево, пользователи смогут оценить взаимосвязи между близкородственными S . mitis групповых видов / штаммов, а также соответствующие им гены вирулентности образуют заметные кластеры на дендрограммах.:max_bytes(150000):strip_icc()/s.pyogenes-5bcb8e9546e0fb0051aacbb8.jpg) Таким образом, этот сравнительный анализ патогеномики может дать отличное представление о профилях генов вирулентности у разных видов Streptococcus . Например, не существует инструмента биоинформатики, который выполняет те же функции, что и PathoProT, а именно прогнозирует и позволяет сравнивать гены вирулентности в геномах разных видов бактерий.
Таким образом, этот сравнительный анализ патогеномики может дать отличное представление о профилях генов вирулентности у разных видов Streptococcus . Например, не существует инструмента биоинформатики, который выполняет те же функции, что и PathoProT, а именно прогнозирует и позволяет сравнивать гены вирулентности в геномах разных видов бактерий. Консервативные гены hasC (hasC1 или SMU . 322c) , которые кодируют UTP-глюкозо-1-фосфат уридилилтрансферазу (или UDP-пирофосфорилазу глюкозы) (M6Spy1871), участвуют в синтезе гиалуроновой кислоты вдоль (HA) капсулы. с двумя соседними генами: hasA и hasB в пределах имеет оперон.[21]. Фактически, это Streptococcus pneumoniae , наиболее патогенный вид S . Группа mitis обладает полисахаридной капсулой, которая способствует бактериальному патогенезу [22]. В Streptococcus HA обнаружен, поскольку материал капсулы стрептококка у некоторых видов является важным фактором вирулентности, эффективно маскируя бактерии от распознавания иммунной системой хозяина [23,24], а также защищая их от реактивных оксидов, выделяемых лейкоцитами [25] ].Кроме того, возможно, что ГК играет важную роль в прикреплении стрептококков группы мититов и колонизации эпителиальных клеток, что приводит к устойчивости бактерий к фагоцитозу макрофагами [26–28].
Консервативные гены hasC (hasC1 или SMU . 322c) , которые кодируют UTP-глюкозо-1-фосфат уридилилтрансферазу (или UDP-пирофосфорилазу глюкозы) (M6Spy1871), участвуют в синтезе гиалуроновой кислоты вдоль (HA) капсулы. с двумя соседними генами: hasA и hasB в пределах имеет оперон.[21]. Фактически, это Streptococcus pneumoniae , наиболее патогенный вид S . Группа mitis обладает полисахаридной капсулой, которая способствует бактериальному патогенезу [22]. В Streptococcus HA обнаружен, поскольку материал капсулы стрептококка у некоторых видов является важным фактором вирулентности, эффективно маскируя бактерии от распознавания иммунной системой хозяина [23,24], а также защищая их от реактивных оксидов, выделяемых лейкоцитами [25] ].Кроме того, возможно, что ГК играет важную роль в прикреплении стрептококков группы мититов и колонизации эпителиальных клеток, что приводит к устойчивости бактерий к фагоцитозу макрофагами [26–28].
 Аналогичным образом, консервативный ген lmb кодирует ламинин-связывающий белок, который впервые был идентифицирован в Streptococcus agalactiae [32]. Практически идентичные адгезины позже были обнаружены как у Streptococcus suis [33], так и у Streptococcus pyogenes [34,35].Было предложено, что адгезины lmb помогают в бактериальном патогенезе посредством инвазии поврежденного эпителия [36]. В целом, многие поверхностные липопротеины и адгезины, которые важны для вирулентности и патогенных инфекций, в S очень консервативны. mitis группа бактерий.
Аналогичным образом, консервативный ген lmb кодирует ламинин-связывающий белок, который впервые был идентифицирован в Streptococcus agalactiae [32]. Практически идентичные адгезины позже были обнаружены как у Streptococcus suis [33], так и у Streptococcus pyogenes [34,35].Было предложено, что адгезины lmb помогают в бактериальном патогенезе посредством инвазии поврежденного эпителия [36]. В целом, многие поверхностные липопротеины и адгезины, которые важны для вирулентности и патогенных инфекций, в S очень консервативны. mitis группа бактерий. mitis , S . infantis , S . tigurinus , S . oralis и S ). Это филогенетическое родство S . mitis групповых видов указывает на близкое родство между видами в M Clade и между видами в S Clade. Интересно, что мы обнаружили, что гены rgp могут использоваться для дифференциации двух разных кладов на тепловой карте. Например, эти маркерные гены присутствуют у всех видов S Clade, но отсутствуют у всех видов M Clade.
mitis , S . infantis , S . tigurinus , S . oralis и S ). Это филогенетическое родство S . mitis групповых видов указывает на близкое родство между видами в M Clade и между видами в S Clade. Интересно, что мы обнаружили, что гены rgp могут использоваться для дифференциации двух разных кладов на тепловой карте. Например, эти маркерные гены присутствуют у всех видов S Clade, но отсутствуют у всех видов M Clade. остаток полученного гликолипидного носителя.И rgpB , и rgpF предположительно должны работать поочередно при удлинении цепи рамнана. Гомологичные рамнозилтрансферазы rgpB и rgpF были обнаружены у Streptococcus thermophilus (STER1436) и Streptococcus gordonii (SGO1022). С другой стороны, гены rgpC и rgpD кодируют предполагаемые переносчики ABC, специфичные для RGP (гомологичный STER1434 в S . thermophilus и гомологичный SGO1024 в S . gordonii ), которые играют роль в экспорте полисахаридов [37]. Ген rgpG (гомолог S . gordonii SGO1723) инициирует синтез RGP путем переноса N -ацетилглюкозамин-1-фосфата на липидный носитель [38].
остаток полученного гликолипидного носителя.И rgpB , и rgpF предположительно должны работать поочередно при удлинении цепи рамнана. Гомологичные рамнозилтрансферазы rgpB и rgpF были обнаружены у Streptococcus thermophilus (STER1436) и Streptococcus gordonii (SGO1022). С другой стороны, гены rgpC и rgpD кодируют предполагаемые переносчики ABC, специфичные для RGP (гомологичный STER1434 в S . thermophilus и гомологичный SGO1024 в S . gordonii ), которые играют роль в экспорте полисахаридов [37]. Ген rgpG (гомолог S . gordonii SGO1723) инициирует синтез RGP путем переноса N -ацетилглюкозамин-1-фосфата на липидный носитель [38]. suis [39]. Индукция инфекционного эндокардита по S . Сообщалось, что mutans запускается генами rgp посредством высвобождения оксида азота [40], агрегации тромбоцитов [41] и придания устойчивости к фагоцитозу полиморфно-ядерными лейкоцитами человека [42]. Следовательно, S Clade S . Виды группы mitis , которые продуцируют эти богатые рамнозой полимеры, могут демонстрировать патогенез, отличный от видов M Clade Streptococcus , с целью установления большей вирулентности и увеличения выживаемости в клетках-хозяевах.Недавнее исследование выявило группу стрептококков Sanguinis в качестве распространенного возбудителя транзиторной бактериемии, которая потенциально может привести к инфекционному эндокардиту. Сообщалось также, что эта группа присутствовала в нескольких случаях вирулентной сепсисной инфекции у пациентов с нейтропенией [43].
suis [39]. Индукция инфекционного эндокардита по S . Сообщалось, что mutans запускается генами rgp посредством высвобождения оксида азота [40], агрегации тромбоцитов [41] и придания устойчивости к фагоцитозу полиморфно-ядерными лейкоцитами человека [42]. Следовательно, S Clade S . Виды группы mitis , которые продуцируют эти богатые рамнозой полимеры, могут демонстрировать патогенез, отличный от видов M Clade Streptococcus , с целью установления большей вирулентности и увеличения выживаемости в клетках-хозяевах.Недавнее исследование выявило группу стрептококков Sanguinis в качестве распространенного возбудителя транзиторной бактериемии, которая потенциально может привести к инфекционному эндокардиту. Сообщалось также, что эта группа присутствовала в нескольких случаях вирулентной сепсисной инфекции у пациентов с нейтропенией [43]. Эти эксклюзивные поиски BLAST функционально основаны на автономном инструменте BLAST [44], загруженном из NCBI. Оба механизма BLAST поддерживают три типа функций BLAST, а именно BLASTN, BLASTP и BLASTX. Пользователи могут определять полноту генома (%) и идентичность генома (%) в формах отправки инструментов BLAST. Эти специализированные инструменты BLAST призваны облегчить пользователям поиск сходства их запрашиваемых последовательностей с последовательностями генома Streptococcus , последовательностями генов (стандартный BLAST), а также с генами вирулентности VFDB (VFDB BLAST), что позволяет пользователям проверить, действительно ли их представляющие интерес гены являются генами потенциальной вирулентности с использованием подхода гомологии последовательностей.
Эти эксклюзивные поиски BLAST функционально основаны на автономном инструменте BLAST [44], загруженном из NCBI. Оба механизма BLAST поддерживают три типа функций BLAST, а именно BLASTN, BLASTP и BLASTX. Пользователи могут определять полноту генома (%) и идентичность генома (%) в формах отправки инструментов BLAST. Эти специализированные инструменты BLAST призваны облегчить пользователям поиск сходства их запрашиваемых последовательностей с последовательностями генома Streptococcus , последовательностями генов (стандартный BLAST), а также с генами вирулентности VFDB (VFDB BLAST), что позволяет пользователям проверить, действительно ли их представляющие интерес гены являются генами потенциальной вирулентности с использованием подхода гомологии последовательностей.
 mitis Виды группы являются важными колонизаторами ротовой полости и иногда связаны с серьезными инфекциями [15]. Кроме того, недавно было высказано предположение, что эти организмы играют важную роль в патогенезе гриппа [8].Таким образом, геномное исследование разнообразных S . mitis Бактерии группы необходимы для понимания того, как эти микроорганизмы переходят от комменсального образа жизни во рту к последующему патогенезу. Однако для широкого спектра S не существует специализированной базы данных генома. mitis групповых геномов для сравнительной геномики. Хотя большинство баз данных по биологическим геномам сосредоточены только на содержании генома и генетической изменчивости, мы определили необходимость создания функциональных биоинформатических инструментов для исследования детерминант вирулентности в геномах посредством сравнительной патогеномики, а также для сравнения содержания генома и генетической изменчивости в пределах S .
mitis Виды группы являются важными колонизаторами ротовой полости и иногда связаны с серьезными инфекциями [15]. Кроме того, недавно было высказано предположение, что эти организмы играют важную роль в патогенезе гриппа [8].Таким образом, геномное исследование разнообразных S . mitis Бактерии группы необходимы для понимания того, как эти микроорганизмы переходят от комменсального образа жизни во рту к последующему патогенезу. Однако для широкого спектра S не существует специализированной базы данных генома. mitis групповых геномов для сравнительной геномики. Хотя большинство баз данных по биологическим геномам сосредоточены только на содержании генома и генетической изменчивости, мы определили необходимость создания функциональных биоинформатических инструментов для исследования детерминант вирулентности в геномах посредством сравнительной патогеномики, а также для сравнения содержания генома и генетической изменчивости в пределах S . mitis группа бактерий.
mitis группа бактерий. В отличие от обычного линейного отображения выравниваний, круговая компоновка показывает взаимосвязь между парами позиций с кариотипами и связями, кодирующими положение, размер и ориентацию связанных геномных элементов.
В отличие от обычного линейного отображения выравниваний, круговая компоновка показывает взаимосвязь между парами позиций с кариотипами и связями, кодирующими положение, размер и ориентацию связанных геномных элементов. Реализация конвейера PGC управляется с помощью сценариев Perl. Этот конвейер производит два типа выходных данных: результаты выравнивания NUCmer и высококачественный график Circos (формат SVG). Пользователи могут бесплатно загрузить эти результаты для публикации или дальнейшего анализа на странице результатов PGC.
Реализация конвейера PGC управляется с помощью сценариев Perl. Этот конвейер производит два типа выходных данных: результаты выравнивания NUCmer и высококачественный график Circos (формат SVG). Пользователи могут бесплатно загрузить эти результаты для публикации или дальнейшего анализа на странице результатов PGC. Кроме того, основанный на Java инструмент сравнения Artemis (ACT) с открытым исходным кодом требует, чтобы пользователи сгенерировали файл сравнения, который идентифицирует области гомологии между сборочным и эталонным геномом с помощью таких программ, как BLASTN, TBLASTX или Mummer, которые должны быть загружены в ACT [18].Сравнительная визуализация ACT выполняется с использованием компонентов Artemis. Напротив, наш инструмент PGC обеспечивает однопоточный процесс попарного выравнивания генома и мгновенное отображение графика Circos сравнительного выравнивания.
Кроме того, основанный на Java инструмент сравнения Artemis (ACT) с открытым исходным кодом требует, чтобы пользователи сгенерировали файл сравнения, который идентифицирует области гомологии между сборочным и эталонным геномом с помощью таких программ, как BLASTN, TBLASTX или Mummer, которые должны быть загружены в ACT [18].Сравнительная визуализация ACT выполняется с использованием компонентов Artemis. Напротив, наш инструмент PGC обеспечивает однопоточный процесс попарного выравнивания генома и мгновенное отображение графика Circos сравнительного выравнивания. A и B выделяют отступы парного сравнения генома между S . mitis B6 и S . mitis 17/34.
A и B выделяют отступы парного сравнения генома между S . mitis B6 и S . mitis 17/34. mitis геномов. Исходя из разрыва, наблюдаемого в (indel «A»), потеря гена произошла около позиции 400 000 п.н.
mitis геномов. Исходя из разрыва, наблюдаемого в (indel «A»), потеря гена произошла около позиции 400 000 п.н. Поэтому мы предлагаем S . mitis B6, возможно, недавно приобрел этот интактный профаг. Графическое изображение интактного профага с различными типами фаг-связанных генов показано на.
Поэтому мы предлагаем S . mitis B6, возможно, недавно приобрел этот интактный профаг. Графическое изображение интактного профага с различными типами фаг-связанных генов показано на. митис B6.
митис B6.  6.3.14)
6.3.14) PathoProT использует автономные инструменты BLAST, загруженные с веб-сайта NCBI. VFDB (версия 2012) в настоящее время содержит набор из 19775 экспериментально подтвержденных генов вирулентности, происходящих от широкого круга различных видов бактерий, что обеспечивает полезный ресурс для поиска гомологии последовательностей. Пользователи могут выбрать список из штаммов Streptococcus для сравнительного анализа и установить пороговое значение, например, идентичность генома и полноту для поиска BLAST через нашу предоставленную онлайн-форму.Параметры по умолчанию для конвейера PathoProT установлены на 50% идентичности последовательностей и 50% полноты последовательности для поиска и идентификации генов ортологичной вирулентности в выбранных геномах Streptococcus . Однако пользователи могут применять желаемые отсечки для поиска гомологии, чтобы достичь оптимальных уровней строгости в своих анализах.
PathoProT использует автономные инструменты BLAST, загруженные с веб-сайта NCBI. VFDB (версия 2012) в настоящее время содержит набор из 19775 экспериментально подтвержденных генов вирулентности, происходящих от широкого круга различных видов бактерий, что обеспечивает полезный ресурс для поиска гомологии последовательностей. Пользователи могут выбрать список из штаммов Streptococcus для сравнительного анализа и установить пороговое значение, например, идентичность генома и полноту для поиска BLAST через нашу предоставленную онлайн-форму.Параметры по умолчанию для конвейера PathoProT установлены на 50% идентичности последовательностей и 50% полноты последовательности для поиска и идентификации генов ортологичной вирулентности в выбранных геномах Streptococcus . Однако пользователи могут применять желаемые отсечки для поиска гомологии, чтобы достичь оптимальных уровней строгости в своих анализах. Собственные сценарии Perl будут обрабатывать выходные данные BLAST (сгенерированные путем поиска этих последовательностей запросов в VFDB) для каждого предсказанного RAST белка (последовательности запроса) в выбранных пользователем геномах и определять предполагаемую вирулентность на основе заданных пользователем параметров.Отфильтрованные результаты BLAST объединяются и организуются в матричную таблицу, содержащую информацию о наличии или отсутствии генов вирулентности (строки) и названия штаммов Streptococcus (столбцы). Наконец, PathoProT передаст и обработает этот вывод с помощью наших собственных сценариев R для иерархической кластеризации (алгоритм полной связи) и создания тепловой карты для визуализации. Штаммы Streptococcus будут отсортированы на основе их профилей генов вирулентности (), и будет построено филогенетическое дерево, пользователи смогут оценить взаимосвязи между близкородственными S . mitis групповых видов / штаммов, а также соответствующие им гены вирулентности образуют заметные кластеры на дендрограммах.
Собственные сценарии Perl будут обрабатывать выходные данные BLAST (сгенерированные путем поиска этих последовательностей запросов в VFDB) для каждого предсказанного RAST белка (последовательности запроса) в выбранных пользователем геномах и определять предполагаемую вирулентность на основе заданных пользователем параметров.Отфильтрованные результаты BLAST объединяются и организуются в матричную таблицу, содержащую информацию о наличии или отсутствии генов вирулентности (строки) и названия штаммов Streptococcus (столбцы). Наконец, PathoProT передаст и обработает этот вывод с помощью наших собственных сценариев R для иерархической кластеризации (алгоритм полной связи) и создания тепловой карты для визуализации. Штаммы Streptococcus будут отсортированы на основе их профилей генов вирулентности (), и будет построено филогенетическое дерево, пользователи смогут оценить взаимосвязи между близкородственными S . mitis групповых видов / штаммов, а также соответствующие им гены вирулентности образуют заметные кластеры на дендрограммах. Таким образом, этот сравнительный анализ патогеномики может дать отличное представление о профилях генов вирулентности у разных видов Streptococcus . Например, не существует инструмента биоинформатики, который выполняет те же функции, что и PathoProT, а именно прогнозирует и позволяет сравнивать гены вирулентности в геномах разных видов бактерий.
Таким образом, этот сравнительный анализ патогеномики может дать отличное представление о профилях генов вирулентности у разных видов Streptococcus . Например, не существует инструмента биоинформатики, который выполняет те же функции, что и PathoProT, а именно прогнозирует и позволяет сравнивать гены вирулентности в геномах разных видов бактерий. Консервативные гены hasC (hasC1 или SMU . 322c) , которые кодируют UTP-глюкозо-1-фосфат уридилилтрансферазу (или UDP-пирофосфорилазу глюкозы) (M6Spy1871), участвуют в синтезе гиалуроновой кислоты вдоль (HA) капсулы. с двумя соседними генами: hasA и hasB в пределах имеет оперон.[21]. Фактически, это Streptococcus pneumoniae , наиболее патогенный вид S . Группа mitis обладает полисахаридной капсулой, которая способствует бактериальному патогенезу [22]. В Streptococcus HA обнаружен, поскольку материал капсулы стрептококка у некоторых видов является важным фактором вирулентности, эффективно маскируя бактерии от распознавания иммунной системой хозяина [23,24], а также защищая их от реактивных оксидов, выделяемых лейкоцитами [25] ].Кроме того, возможно, что ГК играет важную роль в прикреплении стрептококков группы мититов и колонизации эпителиальных клеток, что приводит к устойчивости бактерий к фагоцитозу макрофагами [26–28].
Консервативные гены hasC (hasC1 или SMU . 322c) , которые кодируют UTP-глюкозо-1-фосфат уридилилтрансферазу (или UDP-пирофосфорилазу глюкозы) (M6Spy1871), участвуют в синтезе гиалуроновой кислоты вдоль (HA) капсулы. с двумя соседними генами: hasA и hasB в пределах имеет оперон.[21]. Фактически, это Streptococcus pneumoniae , наиболее патогенный вид S . Группа mitis обладает полисахаридной капсулой, которая способствует бактериальному патогенезу [22]. В Streptococcus HA обнаружен, поскольку материал капсулы стрептококка у некоторых видов является важным фактором вирулентности, эффективно маскируя бактерии от распознавания иммунной системой хозяина [23,24], а также защищая их от реактивных оксидов, выделяемых лейкоцитами [25] ].Кроме того, возможно, что ГК играет важную роль в прикреплении стрептококков группы мититов и колонизации эпителиальных клеток, что приводит к устойчивости бактерий к фагоцитозу макрофагами [26–28].
 Аналогичным образом, консервативный ген lmb кодирует ламинин-связывающий белок, который впервые был идентифицирован в Streptococcus agalactiae [32]. Практически идентичные адгезины позже были обнаружены как у Streptococcus suis [33], так и у Streptococcus pyogenes [34,35].Было предложено, что адгезины lmb помогают в бактериальном патогенезе посредством инвазии поврежденного эпителия [36]. В целом, многие поверхностные липопротеины и адгезины, которые важны для вирулентности и патогенных инфекций, в S очень консервативны. mitis группа бактерий.
Аналогичным образом, консервативный ген lmb кодирует ламинин-связывающий белок, который впервые был идентифицирован в Streptococcus agalactiae [32]. Практически идентичные адгезины позже были обнаружены как у Streptococcus suis [33], так и у Streptococcus pyogenes [34,35].Было предложено, что адгезины lmb помогают в бактериальном патогенезе посредством инвазии поврежденного эпителия [36]. В целом, многие поверхностные липопротеины и адгезины, которые важны для вирулентности и патогенных инфекций, в S очень консервативны. mitis группа бактерий. mitis , S . infantis , S . tigurinus , S . oralis и S ). Это филогенетическое родство S . mitis групповых видов указывает на близкое родство между видами в M Clade и между видами в S Clade. Интересно, что мы обнаружили, что гены rgp могут использоваться для дифференциации двух разных кладов на тепловой карте. Например, эти маркерные гены присутствуют у всех видов S Clade, но отсутствуют у всех видов M Clade.
mitis , S . infantis , S . tigurinus , S . oralis и S ). Это филогенетическое родство S . mitis групповых видов указывает на близкое родство между видами в M Clade и между видами в S Clade. Интересно, что мы обнаружили, что гены rgp могут использоваться для дифференциации двух разных кладов на тепловой карте. Например, эти маркерные гены присутствуют у всех видов S Clade, но отсутствуют у всех видов M Clade. остаток полученного гликолипидного носителя.И rgpB , и rgpF предположительно должны работать поочередно при удлинении цепи рамнана. Гомологичные рамнозилтрансферазы rgpB и rgpF были обнаружены у Streptococcus thermophilus (STER1436) и Streptococcus gordonii (SGO1022). С другой стороны, гены rgpC и rgpD кодируют предполагаемые переносчики ABC, специфичные для RGP (гомологичный STER1434 в S . thermophilus и гомологичный SGO1024 в S . gordonii ), которые играют роль в экспорте полисахаридов [37]. Ген rgpG (гомолог S . gordonii SGO1723) инициирует синтез RGP путем переноса N -ацетилглюкозамин-1-фосфата на липидный носитель [38].
остаток полученного гликолипидного носителя.И rgpB , и rgpF предположительно должны работать поочередно при удлинении цепи рамнана. Гомологичные рамнозилтрансферазы rgpB и rgpF были обнаружены у Streptococcus thermophilus (STER1436) и Streptococcus gordonii (SGO1022). С другой стороны, гены rgpC и rgpD кодируют предполагаемые переносчики ABC, специфичные для RGP (гомологичный STER1434 в S . thermophilus и гомологичный SGO1024 в S . gordonii ), которые играют роль в экспорте полисахаридов [37]. Ген rgpG (гомолог S . gordonii SGO1723) инициирует синтез RGP путем переноса N -ацетилглюкозамин-1-фосфата на липидный носитель [38]. suis [39]. Индукция инфекционного эндокардита по S . Сообщалось, что mutans запускается генами rgp посредством высвобождения оксида азота [40], агрегации тромбоцитов [41] и придания устойчивости к фагоцитозу полиморфно-ядерными лейкоцитами человека [42]. Следовательно, S Clade S . Виды группы mitis , которые продуцируют эти богатые рамнозой полимеры, могут демонстрировать патогенез, отличный от видов M Clade Streptococcus , с целью установления большей вирулентности и увеличения выживаемости в клетках-хозяевах.Недавнее исследование выявило группу стрептококков Sanguinis в качестве распространенного возбудителя транзиторной бактериемии, которая потенциально может привести к инфекционному эндокардиту. Сообщалось также, что эта группа присутствовала в нескольких случаях вирулентной сепсисной инфекции у пациентов с нейтропенией [43].
suis [39]. Индукция инфекционного эндокардита по S . Сообщалось, что mutans запускается генами rgp посредством высвобождения оксида азота [40], агрегации тромбоцитов [41] и придания устойчивости к фагоцитозу полиморфно-ядерными лейкоцитами человека [42]. Следовательно, S Clade S . Виды группы mitis , которые продуцируют эти богатые рамнозой полимеры, могут демонстрировать патогенез, отличный от видов M Clade Streptococcus , с целью установления большей вирулентности и увеличения выживаемости в клетках-хозяевах.Недавнее исследование выявило группу стрептококков Sanguinis в качестве распространенного возбудителя транзиторной бактериемии, которая потенциально может привести к инфекционному эндокардиту. Сообщалось также, что эта группа присутствовала в нескольких случаях вирулентной сепсисной инфекции у пациентов с нейтропенией [43]. Эти эксклюзивные поиски BLAST функционально основаны на автономном инструменте BLAST [44], загруженном из NCBI. Оба механизма BLAST поддерживают три типа функций BLAST, а именно BLASTN, BLASTP и BLASTX. Пользователи могут определять полноту генома (%) и идентичность генома (%) в формах отправки инструментов BLAST. Эти специализированные инструменты BLAST призваны облегчить пользователям поиск сходства их запрашиваемых последовательностей с последовательностями генома Streptococcus , последовательностями генов (стандартный BLAST), а также с генами вирулентности VFDB (VFDB BLAST), что позволяет пользователям проверить, действительно ли их представляющие интерес гены являются генами потенциальной вирулентности с использованием подхода гомологии последовательностей.
Эти эксклюзивные поиски BLAST функционально основаны на автономном инструменте BLAST [44], загруженном из NCBI. Оба механизма BLAST поддерживают три типа функций BLAST, а именно BLASTN, BLASTP и BLASTX. Пользователи могут определять полноту генома (%) и идентичность генома (%) в формах отправки инструментов BLAST. Эти специализированные инструменты BLAST призваны облегчить пользователям поиск сходства их запрашиваемых последовательностей с последовательностями генома Streptococcus , последовательностями генов (стандартный BLAST), а также с генами вирулентности VFDB (VFDB BLAST), что позволяет пользователям проверить, действительно ли их представляющие интерес гены являются генами потенциальной вирулентности с использованием подхода гомологии последовательностей.
 mitis Виды группы являются важными колонизаторами ротовой полости и иногда связаны с серьезными инфекциями [15]. Кроме того, недавно было высказано предположение, что эти организмы играют важную роль в патогенезе гриппа [8].Таким образом, геномное исследование разнообразных S . mitis Бактерии группы необходимы для понимания того, как эти микроорганизмы переходят от комменсального образа жизни во рту к последующему патогенезу. Однако для широкого спектра S не существует специализированной базы данных генома. mitis групповых геномов для сравнительной геномики. Хотя большинство баз данных по биологическим геномам сосредоточены только на содержании генома и генетической изменчивости, мы определили необходимость создания функциональных биоинформатических инструментов для исследования детерминант вирулентности в геномах посредством сравнительной патогеномики, а также для сравнения содержания генома и генетической изменчивости в пределах S .
mitis Виды группы являются важными колонизаторами ротовой полости и иногда связаны с серьезными инфекциями [15]. Кроме того, недавно было высказано предположение, что эти организмы играют важную роль в патогенезе гриппа [8].Таким образом, геномное исследование разнообразных S . mitis Бактерии группы необходимы для понимания того, как эти микроорганизмы переходят от комменсального образа жизни во рту к последующему патогенезу. Однако для широкого спектра S не существует специализированной базы данных генома. mitis групповых геномов для сравнительной геномики. Хотя большинство баз данных по биологическим геномам сосредоточены только на содержании генома и генетической изменчивости, мы определили необходимость создания функциональных биоинформатических инструментов для исследования детерминант вирулентности в геномах посредством сравнительной патогеномики, а также для сравнения содержания генома и генетической изменчивости в пределах S . mitis группа бактерий.
mitis группа бактерий.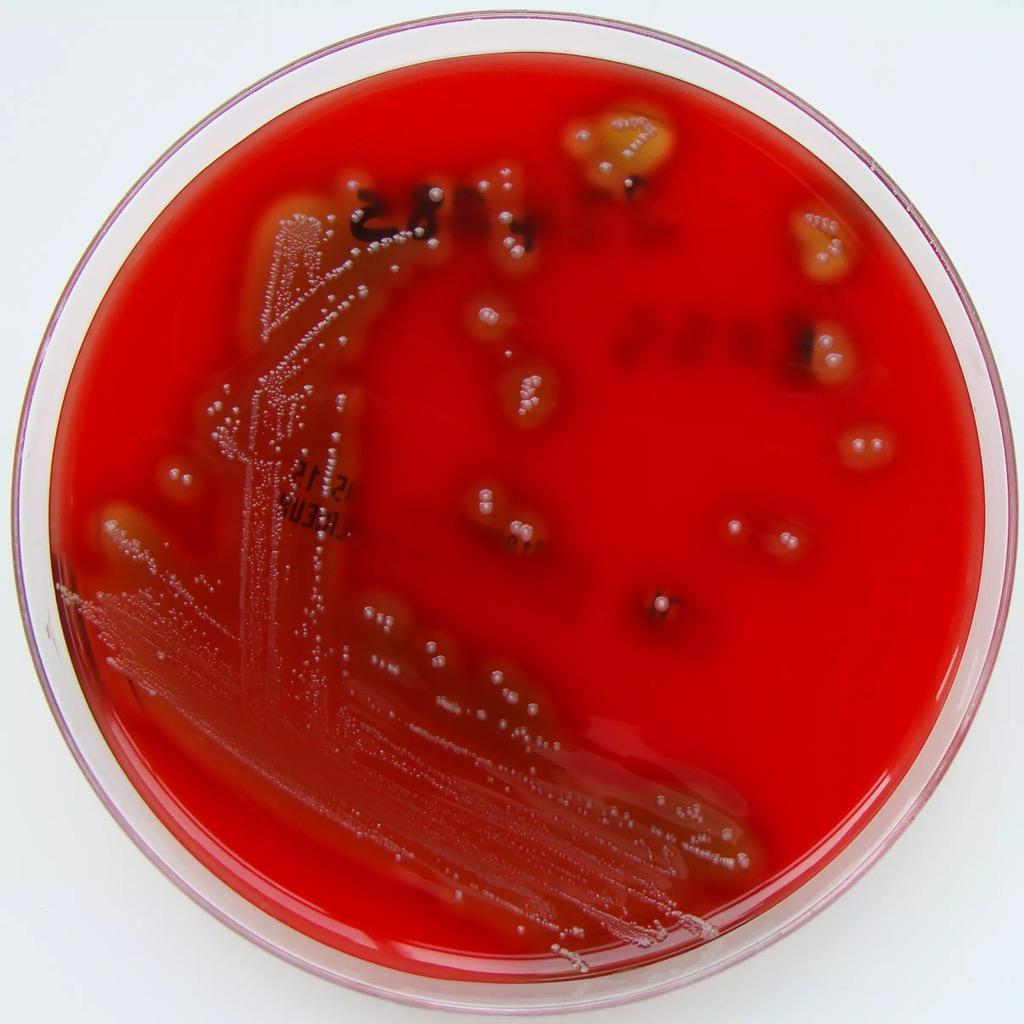 В отличие от обычного линейного отображения выравниваний, круговая компоновка показывает взаимосвязь между парами позиций с кариотипами и связями, кодирующими положение, размер и ориентацию связанных геномных элементов.
В отличие от обычного линейного отображения выравниваний, круговая компоновка показывает взаимосвязь между парами позиций с кариотипами и связями, кодирующими положение, размер и ориентацию связанных геномных элементов. Реализация конвейера PGC управляется с помощью сценариев Perl. Этот конвейер производит два типа выходных данных: результаты выравнивания NUCmer и высококачественный график Circos (формат SVG). Пользователи могут бесплатно загрузить эти результаты для публикации или дальнейшего анализа на странице результатов PGC.
Реализация конвейера PGC управляется с помощью сценариев Perl. Этот конвейер производит два типа выходных данных: результаты выравнивания NUCmer и высококачественный график Circos (формат SVG). Пользователи могут бесплатно загрузить эти результаты для публикации или дальнейшего анализа на странице результатов PGC. Кроме того, основанный на Java инструмент сравнения Artemis (ACT) с открытым исходным кодом требует, чтобы пользователи сгенерировали файл сравнения, который идентифицирует области гомологии между сборочным и эталонным геномом с помощью таких программ, как BLASTN, TBLASTX или Mummer, которые должны быть загружены в ACT [18].Сравнительная визуализация ACT выполняется с использованием компонентов Artemis. Напротив, наш инструмент PGC обеспечивает однопоточный процесс попарного выравнивания генома и мгновенное отображение графика Circos сравнительного выравнивания.
Кроме того, основанный на Java инструмент сравнения Artemis (ACT) с открытым исходным кодом требует, чтобы пользователи сгенерировали файл сравнения, который идентифицирует области гомологии между сборочным и эталонным геномом с помощью таких программ, как BLASTN, TBLASTX или Mummer, которые должны быть загружены в ACT [18].Сравнительная визуализация ACT выполняется с использованием компонентов Artemis. Напротив, наш инструмент PGC обеспечивает однопоточный процесс попарного выравнивания генома и мгновенное отображение графика Circos сравнительного выравнивания. A и B выделяют отступы парного сравнения генома между S . mitis B6 и S . mitis 17/34.
A и B выделяют отступы парного сравнения генома между S . mitis B6 и S . mitis 17/34. mitis геномов. Исходя из разрыва, наблюдаемого в (indel «A»), потеря гена произошла около позиции 400 000 п.н.
mitis геномов. Исходя из разрыва, наблюдаемого в (indel «A»), потеря гена произошла около позиции 400 000 п.н. Поэтому мы предлагаем S . mitis B6, возможно, недавно приобрел этот интактный профаг. Графическое изображение интактного профага с различными типами фаг-связанных генов показано на.
Поэтому мы предлагаем S . mitis B6, возможно, недавно приобрел этот интактный профаг. Графическое изображение интактного профага с различными типами фаг-связанных генов показано на. митис B6.
митис B6.  6.3.14)
6.3.14) PathoProT использует автономные инструменты BLAST, загруженные с веб-сайта NCBI. VFDB (версия 2012) в настоящее время содержит набор из 19775 экспериментально подтвержденных генов вирулентности, происходящих от широкого круга различных видов бактерий, что обеспечивает полезный ресурс для поиска гомологии последовательностей. Пользователи могут выбрать список из штаммов Streptococcus для сравнительного анализа и установить пороговое значение, например, идентичность генома и полноту для поиска BLAST через нашу предоставленную онлайн-форму.Параметры по умолчанию для конвейера PathoProT установлены на 50% идентичности последовательностей и 50% полноты последовательности для поиска и идентификации генов ортологичной вирулентности в выбранных геномах Streptococcus . Однако пользователи могут применять желаемые отсечки для поиска гомологии, чтобы достичь оптимальных уровней строгости в своих анализах.
PathoProT использует автономные инструменты BLAST, загруженные с веб-сайта NCBI. VFDB (версия 2012) в настоящее время содержит набор из 19775 экспериментально подтвержденных генов вирулентности, происходящих от широкого круга различных видов бактерий, что обеспечивает полезный ресурс для поиска гомологии последовательностей. Пользователи могут выбрать список из штаммов Streptococcus для сравнительного анализа и установить пороговое значение, например, идентичность генома и полноту для поиска BLAST через нашу предоставленную онлайн-форму.Параметры по умолчанию для конвейера PathoProT установлены на 50% идентичности последовательностей и 50% полноты последовательности для поиска и идентификации генов ортологичной вирулентности в выбранных геномах Streptococcus . Однако пользователи могут применять желаемые отсечки для поиска гомологии, чтобы достичь оптимальных уровней строгости в своих анализах. Собственные сценарии Perl будут обрабатывать выходные данные BLAST (сгенерированные путем поиска этих последовательностей запросов в VFDB) для каждого предсказанного RAST белка (последовательности запроса) в выбранных пользователем геномах и определять предполагаемую вирулентность на основе заданных пользователем параметров.Отфильтрованные результаты BLAST объединяются и организуются в матричную таблицу, содержащую информацию о наличии или отсутствии генов вирулентности (строки) и названия штаммов Streptococcus (столбцы). Наконец, PathoProT передаст и обработает этот вывод с помощью наших собственных сценариев R для иерархической кластеризации (алгоритм полной связи) и создания тепловой карты для визуализации. Штаммы Streptococcus будут отсортированы на основе их профилей генов вирулентности (), и будет построено филогенетическое дерево, пользователи смогут оценить взаимосвязи между близкородственными S . mitis групповых видов / штаммов, а также соответствующие им гены вирулентности образуют заметные кластеры на дендрограммах.
Собственные сценарии Perl будут обрабатывать выходные данные BLAST (сгенерированные путем поиска этих последовательностей запросов в VFDB) для каждого предсказанного RAST белка (последовательности запроса) в выбранных пользователем геномах и определять предполагаемую вирулентность на основе заданных пользователем параметров.Отфильтрованные результаты BLAST объединяются и организуются в матричную таблицу, содержащую информацию о наличии или отсутствии генов вирулентности (строки) и названия штаммов Streptococcus (столбцы). Наконец, PathoProT передаст и обработает этот вывод с помощью наших собственных сценариев R для иерархической кластеризации (алгоритм полной связи) и создания тепловой карты для визуализации. Штаммы Streptococcus будут отсортированы на основе их профилей генов вирулентности (), и будет построено филогенетическое дерево, пользователи смогут оценить взаимосвязи между близкородственными S . mitis групповых видов / штаммов, а также соответствующие им гены вирулентности образуют заметные кластеры на дендрограммах. Таким образом, этот сравнительный анализ патогеномики может дать отличное представление о профилях генов вирулентности у разных видов Streptococcus . Например, не существует инструмента биоинформатики, который выполняет те же функции, что и PathoProT, а именно прогнозирует и позволяет сравнивать гены вирулентности в геномах разных видов бактерий.
Таким образом, этот сравнительный анализ патогеномики может дать отличное представление о профилях генов вирулентности у разных видов Streptococcus . Например, не существует инструмента биоинформатики, который выполняет те же функции, что и PathoProT, а именно прогнозирует и позволяет сравнивать гены вирулентности в геномах разных видов бактерий. Консервативные гены hasC (hasC1 или SMU . 322c) , которые кодируют UTP-глюкозо-1-фосфат уридилилтрансферазу (или UDP-пирофосфорилазу глюкозы) (M6Spy1871), участвуют в синтезе гиалуроновой кислоты вдоль (HA) капсулы. с двумя соседними генами: hasA и hasB в пределах имеет оперон.[21]. Фактически, это Streptococcus pneumoniae , наиболее патогенный вид S . Группа mitis обладает полисахаридной капсулой, которая способствует бактериальному патогенезу [22]. В Streptococcus HA обнаружен, поскольку материал капсулы стрептококка у некоторых видов является важным фактором вирулентности, эффективно маскируя бактерии от распознавания иммунной системой хозяина [23,24], а также защищая их от реактивных оксидов, выделяемых лейкоцитами [25] ].Кроме того, возможно, что ГК играет важную роль в прикреплении стрептококков группы мититов и колонизации эпителиальных клеток, что приводит к устойчивости бактерий к фагоцитозу макрофагами [26–28].
Консервативные гены hasC (hasC1 или SMU . 322c) , которые кодируют UTP-глюкозо-1-фосфат уридилилтрансферазу (или UDP-пирофосфорилазу глюкозы) (M6Spy1871), участвуют в синтезе гиалуроновой кислоты вдоль (HA) капсулы. с двумя соседними генами: hasA и hasB в пределах имеет оперон.[21]. Фактически, это Streptococcus pneumoniae , наиболее патогенный вид S . Группа mitis обладает полисахаридной капсулой, которая способствует бактериальному патогенезу [22]. В Streptococcus HA обнаружен, поскольку материал капсулы стрептококка у некоторых видов является важным фактором вирулентности, эффективно маскируя бактерии от распознавания иммунной системой хозяина [23,24], а также защищая их от реактивных оксидов, выделяемых лейкоцитами [25] ].Кроме того, возможно, что ГК играет важную роль в прикреплении стрептококков группы мититов и колонизации эпителиальных клеток, что приводит к устойчивости бактерий к фагоцитозу макрофагами [26–28].
 Аналогичным образом, консервативный ген lmb кодирует ламинин-связывающий белок, который впервые был идентифицирован в Streptococcus agalactiae [32]. Практически идентичные адгезины позже были обнаружены как у Streptococcus suis [33], так и у Streptococcus pyogenes [34,35].Было предложено, что адгезины lmb помогают в бактериальном патогенезе посредством инвазии поврежденного эпителия [36]. В целом, многие поверхностные липопротеины и адгезины, которые важны для вирулентности и патогенных инфекций, в S очень консервативны. mitis группа бактерий.
Аналогичным образом, консервативный ген lmb кодирует ламинин-связывающий белок, который впервые был идентифицирован в Streptococcus agalactiae [32]. Практически идентичные адгезины позже были обнаружены как у Streptococcus suis [33], так и у Streptococcus pyogenes [34,35].Было предложено, что адгезины lmb помогают в бактериальном патогенезе посредством инвазии поврежденного эпителия [36]. В целом, многие поверхностные липопротеины и адгезины, которые важны для вирулентности и патогенных инфекций, в S очень консервативны. mitis группа бактерий. mitis , S . infantis , S . tigurinus , S . oralis и S ). Это филогенетическое родство S . mitis групповых видов указывает на близкое родство между видами в M Clade и между видами в S Clade. Интересно, что мы обнаружили, что гены rgp могут использоваться для дифференциации двух разных кладов на тепловой карте. Например, эти маркерные гены присутствуют у всех видов S Clade, но отсутствуют у всех видов M Clade.
mitis , S . infantis , S . tigurinus , S . oralis и S ). Это филогенетическое родство S . mitis групповых видов указывает на близкое родство между видами в M Clade и между видами в S Clade. Интересно, что мы обнаружили, что гены rgp могут использоваться для дифференциации двух разных кладов на тепловой карте. Например, эти маркерные гены присутствуют у всех видов S Clade, но отсутствуют у всех видов M Clade. остаток полученного гликолипидного носителя.И rgpB , и rgpF предположительно должны работать поочередно при удлинении цепи рамнана. Гомологичные рамнозилтрансферазы rgpB и rgpF были обнаружены у Streptococcus thermophilus (STER1436) и Streptococcus gordonii (SGO1022). С другой стороны, гены rgpC и rgpD кодируют предполагаемые переносчики ABC, специфичные для RGP (гомологичный STER1434 в S . thermophilus и гомологичный SGO1024 в S . gordonii ), которые играют роль в экспорте полисахаридов [37]. Ген rgpG (гомолог S . gordonii SGO1723) инициирует синтез RGP путем переноса N -ацетилглюкозамин-1-фосфата на липидный носитель [38].
остаток полученного гликолипидного носителя.И rgpB , и rgpF предположительно должны работать поочередно при удлинении цепи рамнана. Гомологичные рамнозилтрансферазы rgpB и rgpF были обнаружены у Streptococcus thermophilus (STER1436) и Streptococcus gordonii (SGO1022). С другой стороны, гены rgpC и rgpD кодируют предполагаемые переносчики ABC, специфичные для RGP (гомологичный STER1434 в S . thermophilus и гомологичный SGO1024 в S . gordonii ), которые играют роль в экспорте полисахаридов [37]. Ген rgpG (гомолог S . gordonii SGO1723) инициирует синтез RGP путем переноса N -ацетилглюкозамин-1-фосфата на липидный носитель [38]. suis [39]. Индукция инфекционного эндокардита по S . Сообщалось, что mutans запускается генами rgp посредством высвобождения оксида азота [40], агрегации тромбоцитов [41] и придания устойчивости к фагоцитозу полиморфно-ядерными лейкоцитами человека [42]. Следовательно, S Clade S . Виды группы mitis , которые продуцируют эти богатые рамнозой полимеры, могут демонстрировать патогенез, отличный от видов M Clade Streptococcus , с целью установления большей вирулентности и увеличения выживаемости в клетках-хозяевах.Недавнее исследование выявило группу стрептококков Sanguinis в качестве распространенного возбудителя транзиторной бактериемии, которая потенциально может привести к инфекционному эндокардиту. Сообщалось также, что эта группа присутствовала в нескольких случаях вирулентной сепсисной инфекции у пациентов с нейтропенией [43].
suis [39]. Индукция инфекционного эндокардита по S . Сообщалось, что mutans запускается генами rgp посредством высвобождения оксида азота [40], агрегации тромбоцитов [41] и придания устойчивости к фагоцитозу полиморфно-ядерными лейкоцитами человека [42]. Следовательно, S Clade S . Виды группы mitis , которые продуцируют эти богатые рамнозой полимеры, могут демонстрировать патогенез, отличный от видов M Clade Streptococcus , с целью установления большей вирулентности и увеличения выживаемости в клетках-хозяевах.Недавнее исследование выявило группу стрептококков Sanguinis в качестве распространенного возбудителя транзиторной бактериемии, которая потенциально может привести к инфекционному эндокардиту. Сообщалось также, что эта группа присутствовала в нескольких случаях вирулентной сепсисной инфекции у пациентов с нейтропенией [43]. Эти эксклюзивные поиски BLAST функционально основаны на автономном инструменте BLAST [44], загруженном из NCBI. Оба механизма BLAST поддерживают три типа функций BLAST, а именно BLASTN, BLASTP и BLASTX. Пользователи могут определять полноту генома (%) и идентичность генома (%) в формах отправки инструментов BLAST. Эти специализированные инструменты BLAST призваны облегчить пользователям поиск сходства их запрашиваемых последовательностей с последовательностями генома Streptococcus , последовательностями генов (стандартный BLAST), а также с генами вирулентности VFDB (VFDB BLAST), что позволяет пользователям проверить, действительно ли их представляющие интерес гены являются генами потенциальной вирулентности с использованием подхода гомологии последовательностей.
Эти эксклюзивные поиски BLAST функционально основаны на автономном инструменте BLAST [44], загруженном из NCBI. Оба механизма BLAST поддерживают три типа функций BLAST, а именно BLASTN, BLASTP и BLASTX. Пользователи могут определять полноту генома (%) и идентичность генома (%) в формах отправки инструментов BLAST. Эти специализированные инструменты BLAST призваны облегчить пользователям поиск сходства их запрашиваемых последовательностей с последовательностями генома Streptococcus , последовательностями генов (стандартный BLAST), а также с генами вирулентности VFDB (VFDB BLAST), что позволяет пользователям проверить, действительно ли их представляющие интерес гены являются генами потенциальной вирулентности с использованием подхода гомологии последовательностей.
 mitis Виды группы являются важными колонизаторами ротовой полости и иногда связаны с серьезными инфекциями [15]. Кроме того, недавно было высказано предположение, что эти организмы играют важную роль в патогенезе гриппа [8].Таким образом, геномное исследование разнообразных S . mitis Бактерии группы необходимы для понимания того, как эти микроорганизмы переходят от комменсального образа жизни во рту к последующему патогенезу. Однако для широкого спектра S не существует специализированной базы данных генома. mitis групповых геномов для сравнительной геномики. Хотя большинство баз данных по биологическим геномам сосредоточены только на содержании генома и генетической изменчивости, мы определили необходимость создания функциональных биоинформатических инструментов для исследования детерминант вирулентности в геномах посредством сравнительной патогеномики, а также для сравнения содержания генома и генетической изменчивости в пределах S .
mitis Виды группы являются важными колонизаторами ротовой полости и иногда связаны с серьезными инфекциями [15]. Кроме того, недавно было высказано предположение, что эти организмы играют важную роль в патогенезе гриппа [8].Таким образом, геномное исследование разнообразных S . mitis Бактерии группы необходимы для понимания того, как эти микроорганизмы переходят от комменсального образа жизни во рту к последующему патогенезу. Однако для широкого спектра S не существует специализированной базы данных генома. mitis групповых геномов для сравнительной геномики. Хотя большинство баз данных по биологическим геномам сосредоточены только на содержании генома и генетической изменчивости, мы определили необходимость создания функциональных биоинформатических инструментов для исследования детерминант вирулентности в геномах посредством сравнительной патогеномики, а также для сравнения содержания генома и генетической изменчивости в пределах S . mitis группа бактерий.
mitis группа бактерий. В отличие от обычного линейного отображения выравниваний, круговая компоновка показывает взаимосвязь между парами позиций с кариотипами и связями, кодирующими положение, размер и ориентацию связанных геномных элементов.
В отличие от обычного линейного отображения выравниваний, круговая компоновка показывает взаимосвязь между парами позиций с кариотипами и связями, кодирующими положение, размер и ориентацию связанных геномных элементов. Реализация конвейера PGC управляется с помощью сценариев Perl. Этот конвейер производит два типа выходных данных: результаты выравнивания NUCmer и высококачественный график Circos (формат SVG). Пользователи могут бесплатно загрузить эти результаты для публикации или дальнейшего анализа на странице результатов PGC.
Реализация конвейера PGC управляется с помощью сценариев Perl. Этот конвейер производит два типа выходных данных: результаты выравнивания NUCmer и высококачественный график Circos (формат SVG). Пользователи могут бесплатно загрузить эти результаты для публикации или дальнейшего анализа на странице результатов PGC. Кроме того, основанный на Java инструмент сравнения Artemis (ACT) с открытым исходным кодом требует, чтобы пользователи сгенерировали файл сравнения, который идентифицирует области гомологии между сборочным и эталонным геномом с помощью таких программ, как BLASTN, TBLASTX или Mummer, которые должны быть загружены в ACT [18].Сравнительная визуализация ACT выполняется с использованием компонентов Artemis. Напротив, наш инструмент PGC обеспечивает однопоточный процесс попарного выравнивания генома и мгновенное отображение графика Circos сравнительного выравнивания.
Кроме того, основанный на Java инструмент сравнения Artemis (ACT) с открытым исходным кодом требует, чтобы пользователи сгенерировали файл сравнения, который идентифицирует области гомологии между сборочным и эталонным геномом с помощью таких программ, как BLASTN, TBLASTX или Mummer, которые должны быть загружены в ACT [18].Сравнительная визуализация ACT выполняется с использованием компонентов Artemis. Напротив, наш инструмент PGC обеспечивает однопоточный процесс попарного выравнивания генома и мгновенное отображение графика Circos сравнительного выравнивания. A и B выделяют отступы парного сравнения генома между S . mitis B6 и S . mitis 17/34.
A и B выделяют отступы парного сравнения генома между S . mitis B6 и S . mitis 17/34. mitis геномов. Исходя из разрыва, наблюдаемого в (indel «A»), потеря гена произошла около позиции 400 000 п.н.
mitis геномов. Исходя из разрыва, наблюдаемого в (indel «A»), потеря гена произошла около позиции 400 000 п.н. Поэтому мы предлагаем S . mitis B6, возможно, недавно приобрел этот интактный профаг. Графическое изображение интактного профага с различными типами фаг-связанных генов показано на.
Поэтому мы предлагаем S . mitis B6, возможно, недавно приобрел этот интактный профаг. Графическое изображение интактного профага с различными типами фаг-связанных генов показано на. митис B6.
митис B6.  6.3.14)
6.3.14)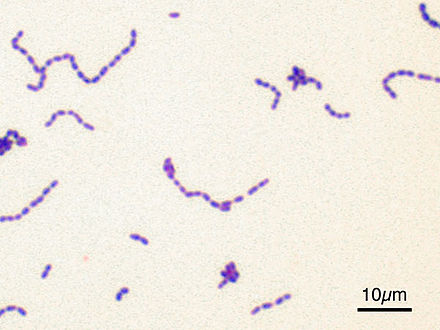 PathoProT использует автономные инструменты BLAST, загруженные с веб-сайта NCBI. VFDB (версия 2012) в настоящее время содержит набор из 19775 экспериментально подтвержденных генов вирулентности, происходящих от широкого круга различных видов бактерий, что обеспечивает полезный ресурс для поиска гомологии последовательностей. Пользователи могут выбрать список из штаммов Streptococcus для сравнительного анализа и установить пороговое значение, например, идентичность генома и полноту для поиска BLAST через нашу предоставленную онлайн-форму.Параметры по умолчанию для конвейера PathoProT установлены на 50% идентичности последовательностей и 50% полноты последовательности для поиска и идентификации генов ортологичной вирулентности в выбранных геномах Streptococcus . Однако пользователи могут применять желаемые отсечки для поиска гомологии, чтобы достичь оптимальных уровней строгости в своих анализах.
PathoProT использует автономные инструменты BLAST, загруженные с веб-сайта NCBI. VFDB (версия 2012) в настоящее время содержит набор из 19775 экспериментально подтвержденных генов вирулентности, происходящих от широкого круга различных видов бактерий, что обеспечивает полезный ресурс для поиска гомологии последовательностей. Пользователи могут выбрать список из штаммов Streptococcus для сравнительного анализа и установить пороговое значение, например, идентичность генома и полноту для поиска BLAST через нашу предоставленную онлайн-форму.Параметры по умолчанию для конвейера PathoProT установлены на 50% идентичности последовательностей и 50% полноты последовательности для поиска и идентификации генов ортологичной вирулентности в выбранных геномах Streptococcus . Однако пользователи могут применять желаемые отсечки для поиска гомологии, чтобы достичь оптимальных уровней строгости в своих анализах.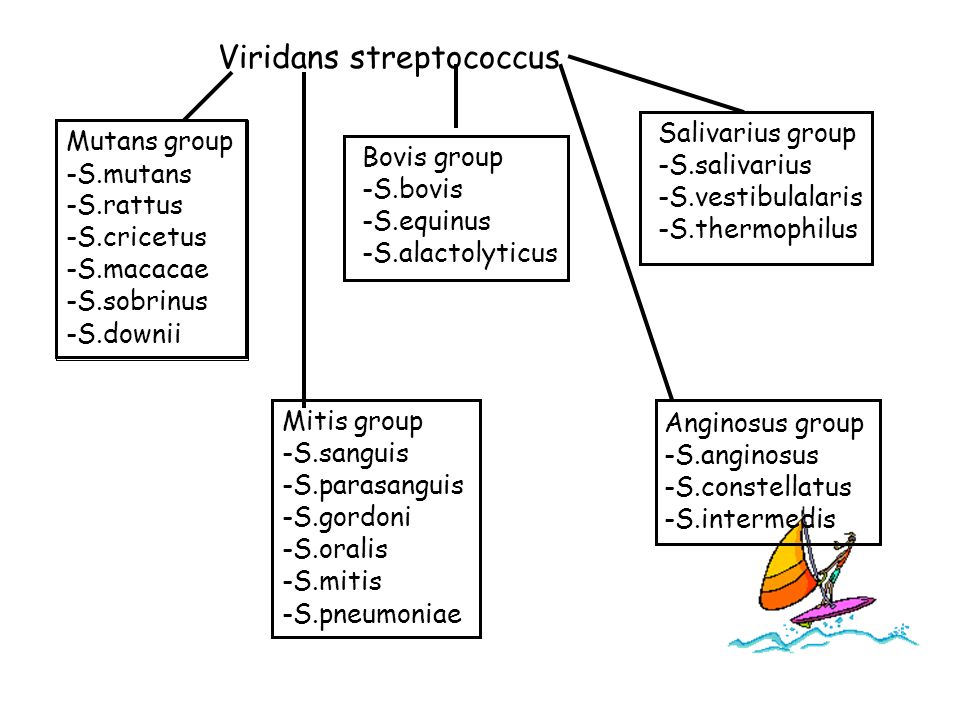 Собственные сценарии Perl будут обрабатывать выходные данные BLAST (сгенерированные путем поиска этих последовательностей запросов в VFDB) для каждого предсказанного RAST белка (последовательности запроса) в выбранных пользователем геномах и определять предполагаемую вирулентность на основе заданных пользователем параметров.Отфильтрованные результаты BLAST объединяются и организуются в матричную таблицу, содержащую информацию о наличии или отсутствии генов вирулентности (строки) и названия штаммов Streptococcus (столбцы). Наконец, PathoProT передаст и обработает этот вывод с помощью наших собственных сценариев R для иерархической кластеризации (алгоритм полной связи) и создания тепловой карты для визуализации. Штаммы Streptococcus будут отсортированы на основе их профилей генов вирулентности (), и будет построено филогенетическое дерево, пользователи смогут оценить взаимосвязи между близкородственными S . mitis групповых видов / штаммов, а также соответствующие им гены вирулентности образуют заметные кластеры на дендрограммах.
Собственные сценарии Perl будут обрабатывать выходные данные BLAST (сгенерированные путем поиска этих последовательностей запросов в VFDB) для каждого предсказанного RAST белка (последовательности запроса) в выбранных пользователем геномах и определять предполагаемую вирулентность на основе заданных пользователем параметров.Отфильтрованные результаты BLAST объединяются и организуются в матричную таблицу, содержащую информацию о наличии или отсутствии генов вирулентности (строки) и названия штаммов Streptococcus (столбцы). Наконец, PathoProT передаст и обработает этот вывод с помощью наших собственных сценариев R для иерархической кластеризации (алгоритм полной связи) и создания тепловой карты для визуализации. Штаммы Streptococcus будут отсортированы на основе их профилей генов вирулентности (), и будет построено филогенетическое дерево, пользователи смогут оценить взаимосвязи между близкородственными S . mitis групповых видов / штаммов, а также соответствующие им гены вирулентности образуют заметные кластеры на дендрограммах. Таким образом, этот сравнительный анализ патогеномики может дать отличное представление о профилях генов вирулентности у разных видов Streptococcus . Например, не существует инструмента биоинформатики, который выполняет те же функции, что и PathoProT, а именно прогнозирует и позволяет сравнивать гены вирулентности в геномах разных видов бактерий.
Таким образом, этот сравнительный анализ патогеномики может дать отличное представление о профилях генов вирулентности у разных видов Streptococcus . Например, не существует инструмента биоинформатики, который выполняет те же функции, что и PathoProT, а именно прогнозирует и позволяет сравнивать гены вирулентности в геномах разных видов бактерий. Консервативные гены hasC (hasC1 или SMU . 322c) , которые кодируют UTP-глюкозо-1-фосфат уридилилтрансферазу (или UDP-пирофосфорилазу глюкозы) (M6Spy1871), участвуют в синтезе гиалуроновой кислоты вдоль (HA) капсулы. с двумя соседними генами: hasA и hasB в пределах имеет оперон.[21]. Фактически, это Streptococcus pneumoniae , наиболее патогенный вид S . Группа mitis обладает полисахаридной капсулой, которая способствует бактериальному патогенезу [22]. В Streptococcus HA обнаружен, поскольку материал капсулы стрептококка у некоторых видов является важным фактором вирулентности, эффективно маскируя бактерии от распознавания иммунной системой хозяина [23,24], а также защищая их от реактивных оксидов, выделяемых лейкоцитами [25] ].Кроме того, возможно, что ГК играет важную роль в прикреплении стрептококков группы мититов и колонизации эпителиальных клеток, что приводит к устойчивости бактерий к фагоцитозу макрофагами [26–28].
Консервативные гены hasC (hasC1 или SMU . 322c) , которые кодируют UTP-глюкозо-1-фосфат уридилилтрансферазу (или UDP-пирофосфорилазу глюкозы) (M6Spy1871), участвуют в синтезе гиалуроновой кислоты вдоль (HA) капсулы. с двумя соседними генами: hasA и hasB в пределах имеет оперон.[21]. Фактически, это Streptococcus pneumoniae , наиболее патогенный вид S . Группа mitis обладает полисахаридной капсулой, которая способствует бактериальному патогенезу [22]. В Streptococcus HA обнаружен, поскольку материал капсулы стрептококка у некоторых видов является важным фактором вирулентности, эффективно маскируя бактерии от распознавания иммунной системой хозяина [23,24], а также защищая их от реактивных оксидов, выделяемых лейкоцитами [25] ].Кроме того, возможно, что ГК играет важную роль в прикреплении стрептококков группы мититов и колонизации эпителиальных клеток, что приводит к устойчивости бактерий к фагоцитозу макрофагами [26–28].
 Аналогичным образом, консервативный ген lmb кодирует ламинин-связывающий белок, который впервые был идентифицирован в Streptococcus agalactiae [32]. Практически идентичные адгезины позже были обнаружены как у Streptococcus suis [33], так и у Streptococcus pyogenes [34,35].Было предложено, что адгезины lmb помогают в бактериальном патогенезе посредством инвазии поврежденного эпителия [36]. В целом, многие поверхностные липопротеины и адгезины, которые важны для вирулентности и патогенных инфекций, в S очень консервативны. mitis группа бактерий.
Аналогичным образом, консервативный ген lmb кодирует ламинин-связывающий белок, который впервые был идентифицирован в Streptococcus agalactiae [32]. Практически идентичные адгезины позже были обнаружены как у Streptococcus suis [33], так и у Streptococcus pyogenes [34,35].Было предложено, что адгезины lmb помогают в бактериальном патогенезе посредством инвазии поврежденного эпителия [36]. В целом, многие поверхностные липопротеины и адгезины, которые важны для вирулентности и патогенных инфекций, в S очень консервативны. mitis группа бактерий. mitis , S . infantis , S . tigurinus , S . oralis и S ). Это филогенетическое родство S . mitis групповых видов указывает на близкое родство между видами в M Clade и между видами в S Clade. Интересно, что мы обнаружили, что гены rgp могут использоваться для дифференциации двух разных кладов на тепловой карте. Например, эти маркерные гены присутствуют у всех видов S Clade, но отсутствуют у всех видов M Clade.
mitis , S . infantis , S . tigurinus , S . oralis и S ). Это филогенетическое родство S . mitis групповых видов указывает на близкое родство между видами в M Clade и между видами в S Clade. Интересно, что мы обнаружили, что гены rgp могут использоваться для дифференциации двух разных кладов на тепловой карте. Например, эти маркерные гены присутствуют у всех видов S Clade, но отсутствуют у всех видов M Clade.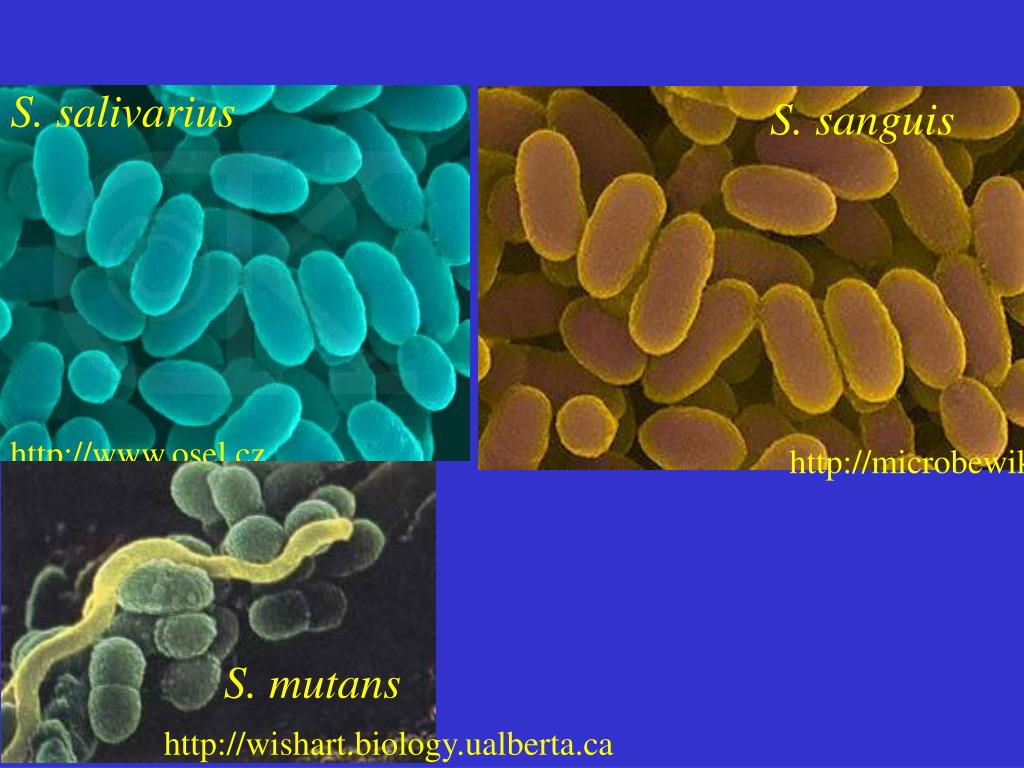 остаток полученного гликолипидного носителя.И rgpB , и rgpF предположительно должны работать поочередно при удлинении цепи рамнана. Гомологичные рамнозилтрансферазы rgpB и rgpF были обнаружены у Streptococcus thermophilus (STER1436) и Streptococcus gordonii (SGO1022). С другой стороны, гены rgpC и rgpD кодируют предполагаемые переносчики ABC, специфичные для RGP (гомологичный STER1434 в S . thermophilus и гомологичный SGO1024 в S . gordonii ), которые играют роль в экспорте полисахаридов [37]. Ген rgpG (гомолог S . gordonii SGO1723) инициирует синтез RGP путем переноса N -ацетилглюкозамин-1-фосфата на липидный носитель [38].
остаток полученного гликолипидного носителя.И rgpB , и rgpF предположительно должны работать поочередно при удлинении цепи рамнана. Гомологичные рамнозилтрансферазы rgpB и rgpF были обнаружены у Streptococcus thermophilus (STER1436) и Streptococcus gordonii (SGO1022). С другой стороны, гены rgpC и rgpD кодируют предполагаемые переносчики ABC, специфичные для RGP (гомологичный STER1434 в S . thermophilus и гомологичный SGO1024 в S . gordonii ), которые играют роль в экспорте полисахаридов [37]. Ген rgpG (гомолог S . gordonii SGO1723) инициирует синтез RGP путем переноса N -ацетилглюкозамин-1-фосфата на липидный носитель [38].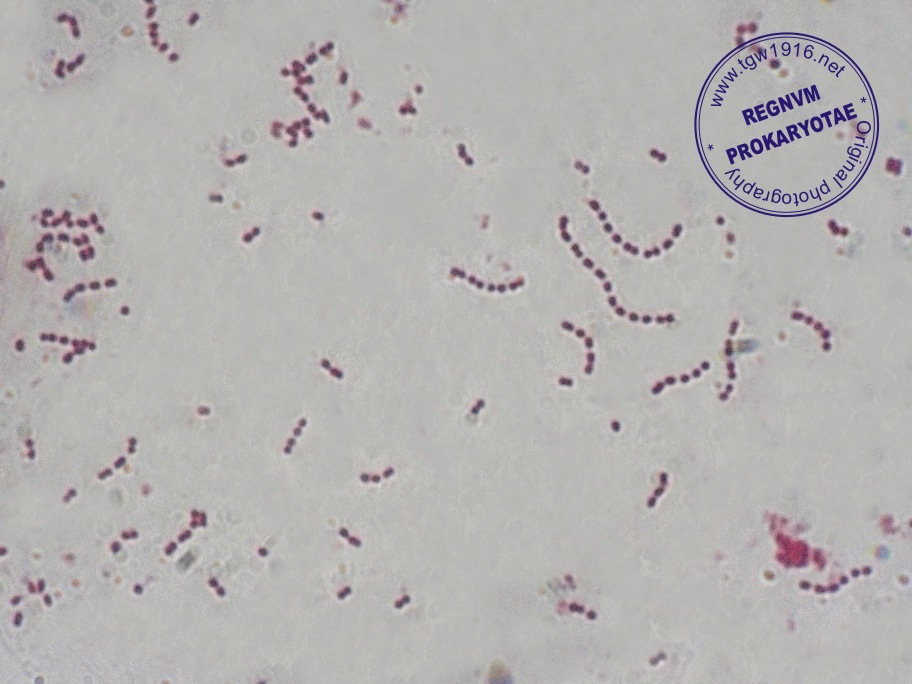 suis [39]. Индукция инфекционного эндокардита по S . Сообщалось, что mutans запускается генами rgp посредством высвобождения оксида азота [40], агрегации тромбоцитов [41] и придания устойчивости к фагоцитозу полиморфно-ядерными лейкоцитами человека [42]. Следовательно, S Clade S . Виды группы mitis , которые продуцируют эти богатые рамнозой полимеры, могут демонстрировать патогенез, отличный от видов M Clade Streptococcus , с целью установления большей вирулентности и увеличения выживаемости в клетках-хозяевах.Недавнее исследование выявило группу стрептококков Sanguinis в качестве распространенного возбудителя транзиторной бактериемии, которая потенциально может привести к инфекционному эндокардиту. Сообщалось также, что эта группа присутствовала в нескольких случаях вирулентной сепсисной инфекции у пациентов с нейтропенией [43].
suis [39]. Индукция инфекционного эндокардита по S . Сообщалось, что mutans запускается генами rgp посредством высвобождения оксида азота [40], агрегации тромбоцитов [41] и придания устойчивости к фагоцитозу полиморфно-ядерными лейкоцитами человека [42]. Следовательно, S Clade S . Виды группы mitis , которые продуцируют эти богатые рамнозой полимеры, могут демонстрировать патогенез, отличный от видов M Clade Streptococcus , с целью установления большей вирулентности и увеличения выживаемости в клетках-хозяевах.Недавнее исследование выявило группу стрептококков Sanguinis в качестве распространенного возбудителя транзиторной бактериемии, которая потенциально может привести к инфекционному эндокардиту. Сообщалось также, что эта группа присутствовала в нескольких случаях вирулентной сепсисной инфекции у пациентов с нейтропенией [43]. Эти эксклюзивные поиски BLAST функционально основаны на автономном инструменте BLAST [44], загруженном из NCBI. Оба механизма BLAST поддерживают три типа функций BLAST, а именно BLASTN, BLASTP и BLASTX. Пользователи могут определять полноту генома (%) и идентичность генома (%) в формах отправки инструментов BLAST. Эти специализированные инструменты BLAST призваны облегчить пользователям поиск сходства их запрашиваемых последовательностей с последовательностями генома Streptococcus , последовательностями генов (стандартный BLAST), а также с генами вирулентности VFDB (VFDB BLAST), что позволяет пользователям проверить, действительно ли их представляющие интерес гены являются генами потенциальной вирулентности с использованием подхода гомологии последовательностей.
Эти эксклюзивные поиски BLAST функционально основаны на автономном инструменте BLAST [44], загруженном из NCBI. Оба механизма BLAST поддерживают три типа функций BLAST, а именно BLASTN, BLASTP и BLASTX. Пользователи могут определять полноту генома (%) и идентичность генома (%) в формах отправки инструментов BLAST. Эти специализированные инструменты BLAST призваны облегчить пользователям поиск сходства их запрашиваемых последовательностей с последовательностями генома Streptococcus , последовательностями генов (стандартный BLAST), а также с генами вирулентности VFDB (VFDB BLAST), что позволяет пользователям проверить, действительно ли их представляющие интерес гены являются генами потенциальной вирулентности с использованием подхода гомологии последовательностей.