
Левомицетин инструкция по применению, цена в аптеках Украины, аналоги, состав, показания | Laevomycetin
Состав
действующее вещество: 1 мл препарата содержит 2,5 мг хлорамфеникола;
вспомогательные вещества: кислота борная, вода для инъекций.
Лекарственная форма
Капли глазные 0,25%.
Основные физико-химические свойства: прозрачная бесцветная жидкость.
Фармакотерапевтическая группа
Средства, применяемые в офтальмологии. Антибиотики. Хлорамфеникол. Код АТХ S01A A01.
Фармакологические свойства
Фармакодинамика.
Левомицетин является антибиотиком широкого спектра действия: эффективен в отношении многих грамположительных и грамотрицательных бактерий, риккетсий, спирохет, возбудителей трахомы, пситтакоза, венерической лимфогранулемы; действует на штаммы бактерий, устойчивых к пенициллину, стрептомицину, сульфаниламидам. Слабоактивен в отношении кислотоустойчивых бактерий, синегнойной палочки, простейших и клостридий.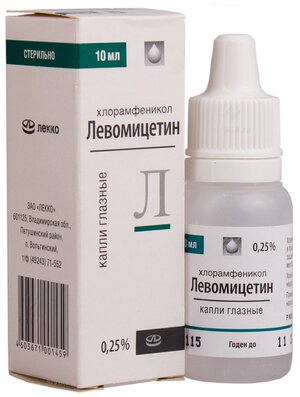
В обычных дозах действует бактериостатически. Механизм антимикробного действия левомицетина связан с нарушением синтеза белков микроорганизмов.
Фармакокинетика.
Препарат хорошо проникает в ткани и жидкости организма; терапевтические концентрации левомицетина при назначении его местно в глазных каплях образуются в стекловидном теле, роговице, радужной оболочке, водянистой влаге глаза; в хрусталик препарат не проникает.
Клинические характеристики
Показания
Конъюнктивиты, кератиты, блефариты, вызванные чувствительными к препарату микроорганизмами.
Противопоказания
Повышенная индивидуальная чувствительность к каким-либо компонентам препарата. Заболевания кожи (псориаз, экзема, грибковые поражения).
Взаимодействие с другими лекарственными средствами и другие виды взаимодействий
Взаимодействие при местном применении глазных капель с другими препаратами не установлено. В случае сопутствующего применения других местных офтальмологических препаратов придерживаться интервала не менее 15 минут между их применением. Глазные мази следует применять последними.
В случае сопутствующего применения других местных офтальмологических препаратов придерживаться интервала не менее 15 минут между их применением. Глазные мази следует применять последними.
Особенности применения.
Не допускается бесконтрольное назначение левомицетина, особенно в педиатрической практике.
Применение в период беременности или кормления грудью.
Препарат противопоказан в период беременности. На период лечения препаратом следует прекратить кормление грудью.
Способность влиять на скорость реакции при управлении автотранспортом или другими механизмами.
В течение 1 часа после приема препарата (пока зрение прояснится) следует воздержаться от управления автотранспортом или работы с другими механизмами.
Способ применения и дозы
Закапывать в конъюнктивальный мешок пипеткой по 1 капле препарата 3 раза в сутки в оба глаза. Курс лечения обычно составляет 5–15 дней (зависит от характера и выраженности заболевания, достигнутого терапевтического эффекта).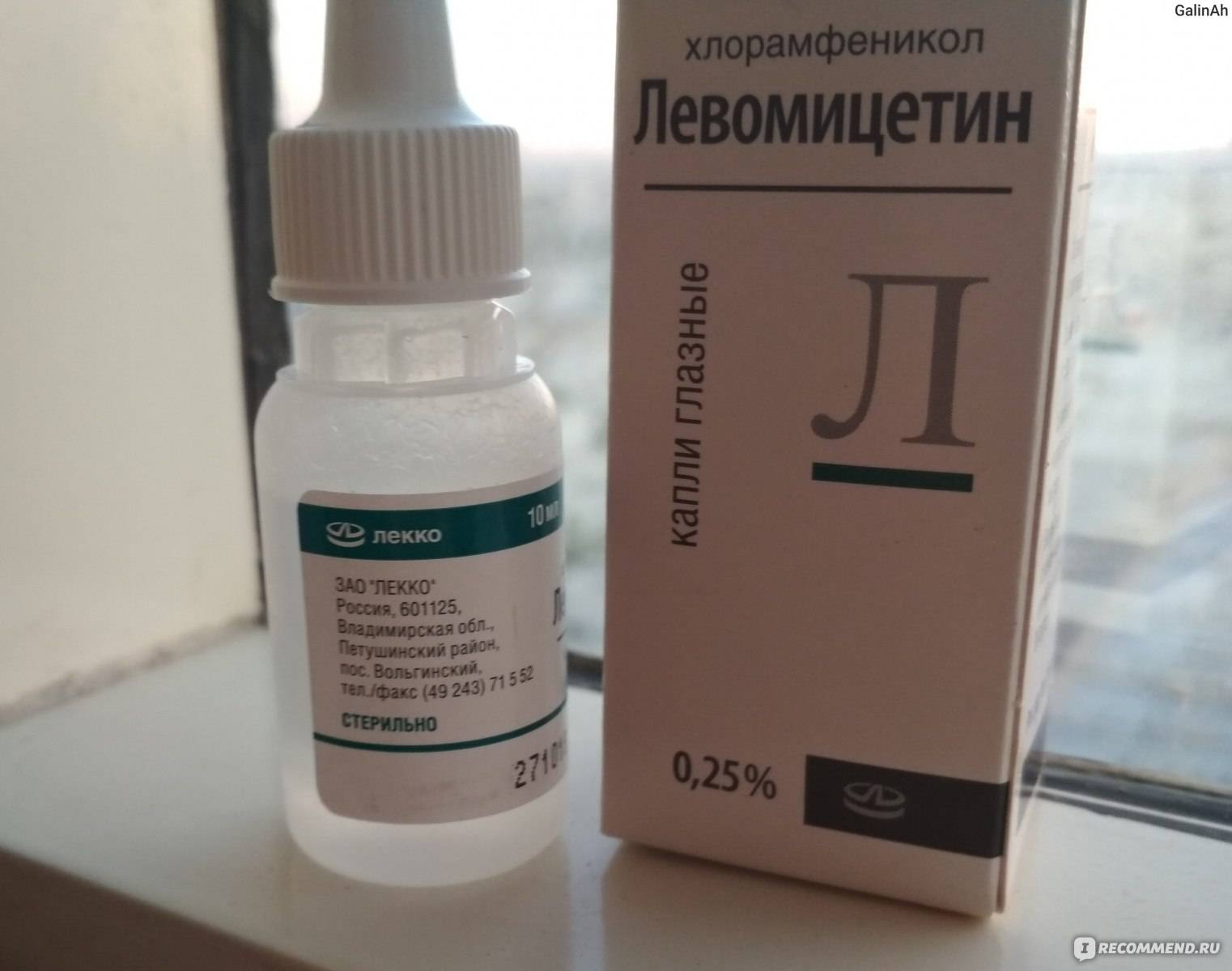
Дети.
Детям до 4 недель назначать только по жизненным показаниям.
Передозировка.
Увеличенные дозы левомицетина могут привести к временному снижению остроты зрения. При таком явлении следует промыть глаза проточной водой.
Побочные реакции
Со стороны иммунной системы, кожи и подкожной клетчатки: аллергические реакции, включая сыпь, ангионевротический отек, крапивницу, зуд, гиперемию кожи.
Со стороны нервной системы: возможна головная боль, головокружение..
Нарушения в месте введения: возможны местные реакции, включая отек век, зуд, раздражение глаз и слезотечение.
Срок годности.
2 года.
После раскрытия флакона — 15 суток.
Не применять после истечения срока годности, указанного на упаковке.
Условия хранения
Хранить в оригинальной упаковке для защиты от воздействия света при температуре не выше 25 оС. Хранить в недоступном для детей месте.
Хранить в недоступном для детей месте.
Упаковка
По 10 мл 0,25% раствора во флаконах пластиковых. По 1 флакону в картонной пачке.
Категория отпуска
По рецепту.
Производитель
ООО «ФЗ «БИОФАРМА», Украина.
Местонахождение производителя и его адрес места осуществления деятельности
Украина, 09100, Киевская область, г. Белая Церковь, ул. Киевская, 37.
Показания к применению
Применяют при ожогах, порезах, трещинах кожи, отитах и заболеваниях, осложненных гнойной инфекцией.
Противопоказания
Повышенная чувствительность к компонентам препарата, дефицит фермента глюкозо-6-фосфатдегидрогеназы, острая интермиттирующая порфирия, кожные заболевания (псориаз, экзема, грибковые поражения), период беременности и грудного вскармливания.
Способ применения
Применяют взрослым и детям старше 1 года. Смазывают пораженные участки кожи (можно применять под окклюзионную повязку). Для лечения ран и ожогов препарат наносят на пораженные участки кожи 4-5 раз в сутки. При гнойных отитах препарат закапывают в наружный слуховой проход (по 2-3 капли). Процедуру повторяют 1 — 2 раза в день, а при значительных выделениях, которые смывают нанесенный раствор, — до 3 — 4 раз в день.
Для лечения ран и ожогов препарат наносят на пораженные участки кожи 4-5 раз в сутки. При гнойных отитах препарат закапывают в наружный слуховой проход (по 2-3 капли). Процедуру повторяют 1 — 2 раза в день, а при значительных выделениях, которые смывают нанесенный раствор, — до 3 — 4 раз в день.
Длительность применения зависит от характера патологического процесса, его выраженности и достигнутого эффекта.
Условия хранения и сроки годности
Хранить при температуре не выше + 25 градусов, в оригинальной упаковке, в защищенном от света и недоступном для детей месте.
Категория отпуска
Без рецепта.
Как избавиться от ячменя быстро
Ячмень считается гнойным острым воспалением сальной железы на веке, возникновение которого связано с проникновением инфекции и ослаблением иммунитета.
Он сопровождается зудом, покраснением века и возникновением боли. Позднее, развивается нарыв и начинает выделяться гной. Поэтому, важно принять срочные меры. Для этого необходимо знать способы как избавиться от ячменя быстро.
Поэтому, важно принять срочные меры. Для этого необходимо знать способы как избавиться от ячменя быстро.
Как избавиться от ячменя быстро методами традиционной медицины
При ячмене, как и при любом воспалительном процессе, эффективным является применение антибиотиков. Рекомендуется закапывать в глаза левомицетин, альбуцид или 1% раствор пенициллина или эритромицина. Делать это нужно до пяти раз в день. Главное, это захватить процесс созревания ячменя, когда он только начинается. При наличии аллергии на один из антибиотиков, необходимо заменить его антибиотиком из другой группы. Для этих целей можно предложить гентамицин или более современные антибиотики такие, как тобрекс или ципролет.
Как избавиться от ячменя?
Способ, который помогает решить проблему как избавиться от ячменя быстро, заключается в использовании тетрациклиновой или эритромициновой мази. Мазь закладывают на ночь за край нижнего века, выполняя следующие действия: левой рукой слегка оттягивается вниз нижнее веко, а правой рукой выдавливается и аккуратно закладывается около трех миллиметров мази.
Как избавиться от ячменя быстро народными средствами?
К одному из способов как избавиться от ячменя быстро относится его прогревание, но только когда он уже вскрылся. Сваренное вкрутую яйцо надо обернуть тонкой мягкой тряпочкой и прикладывать к больному месту до 15 минут. В течение дня делать это нужно 4-5 раз. Слабый раствор ромашки или борной кислоты можно использовать в качестве согревающего компресса, который ускоряет процесс рассасывания ячменя, а также его созревания и выделения гноя.
Примочки и компрессы можно сделать из настоя цветков календулы. Для их приготовления берется 10-15 сухих цветков, которые заливаются одним стаканом кипятка, настаиваются около 40 минут и процеживаются. Также можно делать примочки из сока алоэ. Необходимо срезать лист, промыть, выдавить сок и развести кипяченной водой в пропорции 1: 10. Примочки надо делать до трех раз в день.
Выдавливать гной из ячменя нельзя. Это может стать причиной распространения инфекции по кровеносным сосудам и вызвать сепсис или менингит. В случае необходимости надо обязательно обратиться за консультацией к офтальмологу если у вас ячмень или заворот нижнего века.
В случае необходимости надо обязательно обратиться за консультацией к офтальмологу если у вас ячмень или заворот нижнего века.
быстрое лечение в домашних условиях
Ячменем называется заболевание глазного века, которое характеризуется воспалением в волосяном мешочке ресницы либо в сальной железе около ее луковицы. Медицинское наименование патологии – гордеолум. Оно вызывает дискомфорт, поскольку отекший глаз с ячменем выглядит непривлекательно и сопровождается болевыми ощущениями при движениях верхнего либо нижнего века. Патология может проявиться независимо от возраста – как у взрослого, так и у ребенка. Заболевание поражает обычно один глаз.
Появление ячменя на глазу происходит неожиданно. Когда воспалительный процесс развивается стремительно, больных интересует, что делать в такой ситуации. Требуется немедленное лечение. Нельзя относиться к этому нарушению как к временному неудобству. Его возникновение свидетельствует о том, что иммунные силы организма снижены.
Причины и симптомы
Гордеолум возникает вследствие бактериальной инфекции. В большинстве случаев это золотистый стафилококк – грамположительная бактерия, способная сохраняться на кожных покровах и слизистых.
Провоцируют появление ячменя на глазу разные факторы:
- переохлаждение организма, ослабленный иммунитет, авитаминоз;
- несоблюдение правил гигиены глаз и век;
- стрессовые состояния, отсутствие полноценного отдыха, здорового сна;
- инфекционные заболевания либо период восстановления после них;
- наличие других проблем с органами зрения – конъюнктивита, блефаритов;
- повышенный уровень липидов в крови.
Применение некачественной косметики или ношение контактных линз также могут стать провоцирующими нарушение факторами, но они не являются первопричиной патологического процесса.
Основная симптоматика патологии:
- зуд и жжение века, повышенная слезоточивость;
- ощущение инородного предмета под глазом;
- покраснение, вздутие, отвердение нижнего или верхнего века;
- боль при прикосновении к поврежденной области и при моргании;
- гной в виде головки желтоватого цвета на припухлости;
- иногда – воспаление ближайших лимфатических узлов, повышение температуры.

Заразен ли ячмень
Для окружающих он не опасен, но только при условии соблюдений элементарных гигиенических правил. Риск заражения обычно возникает у маленьких детей. Ввиду своего возраста они еще не понимают важности соблюдения гигиены и поэтому часто страдают бактериальными инфекциями. Заражение возможно и при применении косметики больного. Но в целом изоляция взрослого либо ребенка, страдающего гордеолумом, не требуется.
Возбудители болезни находятся в гное, расположенном в мешочке ячменя. На поверхности бактерии отсутствуют. Но гноя внутри обычно немного, поэтому даже лопнувший ячмень не опасен. В 80% случаев гнойник прорывается ночью, когда больной спит. Гнойные массы, если и попадают куда-либо, то на лицо человека, его подушку, постельное белье. Дальше инфекция не распространяется.
Виды и стадии развития гордеолума
По локализации поражения могут быть следующих видов:
- Наружный ячмень. Более распространен по статистике. Инфекционно-воспалительный процесс локализован на краю века, так как инфекция оседает в сальной железе Цейса либо в ресничной луковице.
- Внутренний ячмень на глазу. Имеет другое название – мейбомит. Обусловлен проникновением патогенных микроорганизмов в проход мейбомиевых желез, находящихся с тыльной стороны края век.
 Более распространен по статистике. Инфекционно-воспалительный процесс локализован на краю века, так как инфекция оседает в сальной железе Цейса либо в ресничной луковице.
Более распространен по статистике. Инфекционно-воспалительный процесс локализован на краю века, так как инфекция оседает в сальной железе Цейса либо в ресничной луковице.По типу выделяют:
- Горячий ячмень. Ему присуще классическое развитие болезни, инфицирование локализуется в ресничном кармане – луковице либо железе Цейса. При терапии проходит примерно за неделю.
- Холодный ячмень (халязион, киста мейбомиева). Воспалительный процесс поражает мейбомиевые железы. Развивается очень медленно. На восстановление уходит 1–2 месяца. При наличии очень большого ячменя возможно его оперативное удаление.
Воспаление проходит стадии:
- Формирование гнойного стержня. Небольшая красноватая припухлость вызывает неприятные ощущения при движении веками.
- Формирование гнойника. Состояние больного на этом этапе могут облегчить лишь медикаменты.
- Прорыв гноя. Не свидетельствует о выздоровлении, однако больному становится гораздо легче. В этот период нужно держать глаза в чистоте и использовать назначенные лекарства, мази.
Как быстро вылечить ячмень
При возникновении ячменя на глазу как лечить его – зависит от этапа развития инфекции. Если принять меры на начальной стадии, есть вероятность того, что воспаление не распространится и отек пройдет через 1–2 суток. Для этого:
- Обрабатывают проблемный участок антисептиком. Подойдет медицинский спирт, который разбавляют с водой в соотношении 1:1, раствор бриллиантового зеленого или йода, настойка календулы.
- Выполняют сухое прогревание. Для этого разогревают на сковороде морскую соль либо любую крупу. Можно взять сваренное вкрутую яйцо. Выбранный источник тепла оборачивают чистой тканью и прикладывают к воспаленному месту, пока компресс не остынет.
Если воспаление все же развилось, лучше обратиться к врачу, поскольку необходимо лечение ячменя медикаментами. Из пораженной области обязательно должны выйти гнойные массы. Если этого не произошло естественным путем, выдавливать гнойник нельзя, чтобы не заразить кровь и другие органы. Офтальмолог в таких случаях делает прокол в стерильных условиях. Шрама от него не остается.
Для уничтожения бактерий используют капли от ячменя у взрослых – Тобрекс, Альбуцид, Левомицетин, Вигамокс, Гентамицин. Их закапывают 3–5 раз в сутки. Важно не допускать прикосновения флакона к глазу.
Быстрое лечение ячменя в домашних условиях обеспечено благодаря сочетанию капель с антибактериальными мазями. Последние закладывают на ночь, поскольку они могут временно снижать зрение. Но при их приобретении в аптеке непременно уточните, что вам нужны препараты именно для глаз. Многие из этих лекарств выпускают также и для кожи, содержание активных веществ в них выше, что может навредить зрительному аппарату. К ним относят Тетрациклин, Синтомицин, Гидрокортизон, Эритромицин, Макситрол.
К ним относят Тетрациклин, Синтомицин, Гидрокортизон, Эритромицин, Макситрол.
Сколько проходит ячмень – зависит от разновидности патологии. Внутренний приходится лечить дольше, чем наружный – на полное выздоровление требуется от 1 месяца.
Профилактические меры
Профилактика включает соблюдение рекомендаций:
- Укрепление иммунных сил. Делать это следует всегда, не только при угрозе гордеолума или в межсезонье. Рекомендуется пить витамины, отдавать предпочтение свежим овощам и фруктам, больше двигаться, бывать на свежем воздухе, соблюдать режим дня.
- Своевременное устранение офтальмологических нарушений. При наличии склонности к глазным заболеваниям нужно регулярно посещать офтальмолога. Самодиагностика и самолечение обычно способствуют переходу патологии в хроническую форму.
- Соблюдение гигиенических норм. Нужно иметь свое личное полотенце, мыть руки с мылом, менять постельное белье каждые 1–2 недели, не пользоваться чужими косметическими средствами.
- Выработка привычки не касаться глаз. Делать этого не стоит даже в обычном состоянии, когда человека не беспокоят никакие проблемы. Это правило распространяется и на лицо – не следует его трогать в течение дня немытыми руками.

Несложные меры профилактики позволят избежать воспаления и сохранить глаза здоровыми.
| Действующее вещество: Тобрамицин | ||
| Тобрекс (капли глазные) (Алкон Фармасьютикалз) |
141,37–250 |
В состав входит современный антибиотик широкого спектра действия из группы аминогликозидов. Применяется при блефарите, конъюнктивите, кератите, иридоциклите и для профилактики послеоперационных осложнений в офтальмологии. |
| Действующее вещество: Ципрофлоксацин | ||
| Офтоципро (Татхимфармпрепараты) Ципролет (капли глазные) (Доктор Редди’с) Ципромед (капли глазные) (Промед Экспортс) Ципрофлоксацин (капли глазные) (Ромфарм) Ципрофлоксацин-АКОС (Синтез) |
125–224 50,5–70 105–143
15–43 12–22 |
Содержит современный антибиотик из группы фторхинолонов. Применяется для лечения блефарита, конъюнктивита, кератита, бактериальных язв роговицы, хронического дакриоцистита (воспаления слезного мешочка) и ячменя. Также используется для предоперационной профилактики в офтальмохирургии и лечения послеоперационных инфекционных осложнений, а также для лечения и профилактики инфекционных осложнений глаз после травм или попадания инородных тел. Иногда может вызывать светобоязнь, слезотечение, ощущение инородного тела в глазах, неприятный привкус во рту сразу после закапывания и ряд других побочных эффектов. Противопоказан при вирусном кератите, беременности, кормлении грудью, детям до 1 года. Применяется для лечения блефарита, конъюнктивита, кератита, бактериальных язв роговицы, хронического дакриоцистита (воспаления слезного мешочка) и ячменя. Также используется для предоперационной профилактики в офтальмохирургии и лечения послеоперационных инфекционных осложнений, а также для лечения и профилактики инфекционных осложнений глаз после травм или попадания инородных тел. Иногда может вызывать светобоязнь, слезотечение, ощущение инородного тела в глазах, неприятный привкус во рту сразу после закапывания и ряд других побочных эффектов. Противопоказан при вирусном кератите, беременности, кормлении грудью, детям до 1 года. |
| Действующее вещество: Офлоксацин | ||
| Данцил (капли глазные/ушные) (Промед Экспортс) Офлоксацин (мазь глазная) (Синтез) Флоксал (мазь глазная, капли глазные) (Бауш энд Ломб) |
128,5–194,5
29,1–65 184–406 |
В состав препарата входит антибиотик широкого спектра действия из группы фторхинолонов. Применяется при бактериальных заболеваниях век, конъюнктивы и роговицы, ячмене, дакриоцистите, хламидийных инфекциях глаз, а также для профилактики инфекционных осложнений после хирургических вмешательств по поводу удаления инородного тела и травмы глаза. Может вызывать ощущение жжения и дискомфорта в глазах, покраснение, зуд и сухость конъюнктивы, светобоязнь, слезотечение. В большинстве случаев эти симптомы непродолжительные. Противопоказан при хроническом небактериальном конъюнктивите, беременности, кормлении грудью и детям до 15 лет. Применяется при бактериальных заболеваниях век, конъюнктивы и роговицы, ячмене, дакриоцистите, хламидийных инфекциях глаз, а также для профилактики инфекционных осложнений после хирургических вмешательств по поводу удаления инородного тела и травмы глаза. Может вызывать ощущение жжения и дискомфорта в глазах, покраснение, зуд и сухость конъюнктивы, светобоязнь, слезотечение. В большинстве случаев эти симптомы непродолжительные. Противопоказан при хроническом небактериальном конъюнктивите, беременности, кормлении грудью и детям до 15 лет. |
| Действующее вещество: Левофлоксацин | ||
| Сигницеф (капли глазные) (Промед Экспортс) |
150–275 |
Антибактериальный препарат группы фторхинолонов. Применяется для лечения конъюнктивита, блефарита, кератита и других заболеваний глаз, вызванных чувствительными к левофлоксацину бактериями, а также для профилактики осложнений после хирургических и лазерных операций на глазу. Иногда может вызывать снижение остроты зрения и появление слизистых тяжей, неприятные ощущения в глазу, синдром сухого глаза и ряд других побочных эффектов. Противопоказан при беременности, кормлении грудью, детям до 1 года. Иногда может вызывать снижение остроты зрения и появление слизистых тяжей, неприятные ощущения в глазу, синдром сухого глаза и ряд других побочных эффектов. Противопоказан при беременности, кормлении грудью, детям до 1 года. |
| Действующее вещество: Эритромицин | ||
| Эритромицин (мазь глазная) (разные производители) |
15–50 |
В состав включены антибиотики широкого спектра действия. Применяются при конъюнктивите, блефарите, кератите и других заболеваниях глаз. Используются уже несколько десятилетий, поэтому иногда могут действовать недостаточно эффективно из-за того, что часть болезнетворных бактерий выработали к ним устойчивость. |
| Действующее вещество: Тетрациклин | ||
| Тетрациклиновая глазная мазь (Татхимфармпрепараты) |
29–44,4 |
В состав включены антибиотики широкого спектра действия. Применяются при конъюнктивите, блефарите, кератите и других заболеваниях глаз. Используются уже несколько десятилетий, поэтому иногда могут действовать недостаточно эффективно из-за того, что часть болезнетворных бактерий выработали к ним устойчивость. Применяются при конъюнктивите, блефарите, кератите и других заболеваниях глаз. Используются уже несколько десятилетий, поэтому иногда могут действовать недостаточно эффективно из-за того, что часть болезнетворных бактерий выработали к ним устойчивость. |
| Действующее вещество: Хлорамфеникол | ||
| Левомицетин (капли глазные) (разные производители) |
7–34 |
В состав включены антибиотики широкого спектра действия. Применяются при конъюнктивите, блефарите, кератите и других заболеваниях глаз. Используются уже несколько десятилетий, поэтому иногда могут действовать недостаточно эффективно из-за того, что часть болезнетворных бактерий выработали к ним устойчивость. |
| Действующее вещество: Норфлоксацин | ||
| Нормакс (капли глазные/ушные) (Ипка) |
73,5–139 |
Антибактериальный препарат из группы фторхинолонов. Применяется для лечения конъюнктивита, блефарита, кератита и других заболеваний глаз, а также при отитах. Используется для профилактики глазных инфекций после удаления инородного тела из роговицы или конъюнктивы, после повреждения химическими или физическими средствами, до и после хирургических вмешательств. Иногда может вызывать тошноту, изжогу, головную боль, головокружение, чувство усталости и другие побочные эффекты. Не рекомендовано назначение препарата при беременности, кормлении грудью, а также детям и подросткам в возрасте до 18 лет. Применяется для лечения конъюнктивита, блефарита, кератита и других заболеваний глаз, а также при отитах. Используется для профилактики глазных инфекций после удаления инородного тела из роговицы или конъюнктивы, после повреждения химическими или физическими средствами, до и после хирургических вмешательств. Иногда может вызывать тошноту, изжогу, головную боль, головокружение, чувство усталости и другие побочные эффекты. Не рекомендовано назначение препарата при беременности, кормлении грудью, а также детям и подросткам в возрасте до 18 лет. |
| Действующее вещество: Фузидовая кислота | ||
| Фуциталмик (капли глазные) (Лео Фармасьютикал) |
245–503 |
Антибиотик из группы фузидинов. Применяется при конъюнктивите, блефарите, кератите и дакриоцистите. Может вызывать жжение, зуд, ощущение сухости и болезненности в глазу, кратковременное затуманивание зрения и другие побочные эффекты. |
| Действующее вещество: Пиклоксидин | ||
| Витабакт (капли глазные) (Лаборатуар Теа) |
225–342 |
Препарат с противомикробным действием. Применяется у взрослых и детей с рождения при бактериальных инфекциях глаз, дакриоцистите, а также для профилактики инфекционных осложнений после хирургических вмешательств. |
| Действующее вещество: Сульфацетамид | ||
| Сульфацил-натрия (капли глазные) (разные производители) |
32–99 |
Антибактериальное средство из группы сульфаниламидов. Применяется при гнойных язвах роговицы, конъюнктивитах, блефаритах, гонорейных заболеваниях глаз у новорожденных и взрослых. Может вызвать зуд, покраснение, отечность. Может вызвать зуд, покраснение, отечность. |
| Действующее вещество: Цинка сульфат + борная кислота | ||
| Цинка сульфат и борная кислота (капли глазные) (Обновление, Диафарм) |
7–15 |
Комбинированный препарат. Цинка сульфат оказывает противомикробное, вяжущее, подсушивающее и местное противовоспалительное действие. Борная кислота – антисептическое средство. Применяется при конъюнктивите. |
| Действующее вещество: Гентамицин + дексаметазон | ||
| Декса-гентамицин (капли глазные) (Урсафарм Арцнаймиттель) |
66–127 |
Комбинированный препарат для местного применения в офтальмологии. Оказывает противоаллергическое, противовоспалительное и антибактериальное действие. Применяется при конъюнктивите, кератите, блефарите, ячмене, аллергии на глазах и веках, сопровождающейся бактериальным инфицированием, а также для профилактики и лечения воспалительных явлений в послеоперационном периоде. Может вызывать ощущение жжения в глазу после применения препарата, грибковые инфекции и другие побочные эффекты. Применение после травм роговицы может приводить к замедлению ее заживления. Противопоказан при герпетическом кератите и других вирусных заболеваниях роговицы и конъюнктивы, а также при грибковых и туберкулезных заболеваниях глаз, повышенном внутриглазном давлении и ряде других состояний. Не рекомендовано применение в I триместре беременности, детям и подросткам до 18 лет и при ношении контактных линз. Применяется при конъюнктивите, кератите, блефарите, ячмене, аллергии на глазах и веках, сопровождающейся бактериальным инфицированием, а также для профилактики и лечения воспалительных явлений в послеоперационном периоде. Может вызывать ощущение жжения в глазу после применения препарата, грибковые инфекции и другие побочные эффекты. Применение после травм роговицы может приводить к замедлению ее заживления. Противопоказан при герпетическом кератите и других вирусных заболеваниях роговицы и конъюнктивы, а также при грибковых и туберкулезных заболеваниях глаз, повышенном внутриглазном давлении и ряде других состояний. Не рекомендовано применение в I триместре беременности, детям и подросткам до 18 лет и при ношении контактных линз. |
| Действующее вещество: Тобрамицин + дексаметазон | ||
| Тобрадекс (капли глазные) (Алкон Фармасьютикалз) Тобразон (капли глазные) (Кадила Фармасьютикалз) |
215–396,85
80–185 |
Комбинированный препарат с антибактериальным и противовоспалительным действием. Применяется для профилактики послеоперационных инфекционных осложнений, блефарите, конъюнктивите, кератите без повреждения эпителия. Может вызывать повышение внутриглазного давления, развитие вторичной инфекции как следствие подавления защитной реакции организма пациента. Противопоказан при вирусных заболеваниях роговицы и конъюнктивы, туберкулезных и грибковых заболеваниях глаз, после удаления инородного тела роговицы. |
| Действующее вещество: Неомицин + полимиксин В + дексаметазон | ||
| Макситрол (капли глазные) (Алкон Фармасьютикалз) |
265–532 |
Комбинированный препарат с антибактериальным и противовоспалительным действием. Применяется при блефарите, конъюнктивите, кератите, иридоциклите (воспалении радужной оболочки), а также для профилактики послеоперационных инфекционных осложнений. Может вызывать повышение внутриглазного давления, замедление процесса заживления ран и другие побочные эффекты. Противопоказания – как у тобрамицина в сочетании с дексаметазоном. Может вызывать повышение внутриглазного давления, замедление процесса заживления ран и другие побочные эффекты. Противопоказания – как у тобрамицина в сочетании с дексаметазоном. |
левомицетин капли ребенку — 25 рекомендаций на Babyblog.ru
стащила!!!!!!!!!!!!(как лечить ребенка)
27 сентября, 11:52
ЕСЛИ ТЕМПЕРАТУРА 38,3-38,8
-Нурофен (сироп/свечи)или
-Эффералган (сироп/свечи)или
-Панадол(свечи)-сироп не покупать, очень аллергенный или
-Цифекон(свечи)
Купить свечи и сироп из списка,их можно чередовать при повторном повышении температуры через 3,5-4 часа.
ЕСЛИ ТЕМПЕРАТУРА 38,8-39,2
-Анальгиновые свечи 0,25
ЕСЛИ ТЕМПЕРАТУРА больше 39,2
-1/3 таб. анальгина+1/3 таб. парацетамола(0,5 г)+1/3 таб. супрастина
и/или 1/3 таб. но-шпы (40 мг). Все смешать.
Раздеть, обтирать теплой водкой, но не кисти рук и не стопы.
• ***ПРОТИВОВИРУСНЫЕ***
-Оцилококкцинум по 1 контейнеру х 2 раза в день 3 дня- начало заболевания
или как профилактика, если кто-то заболел.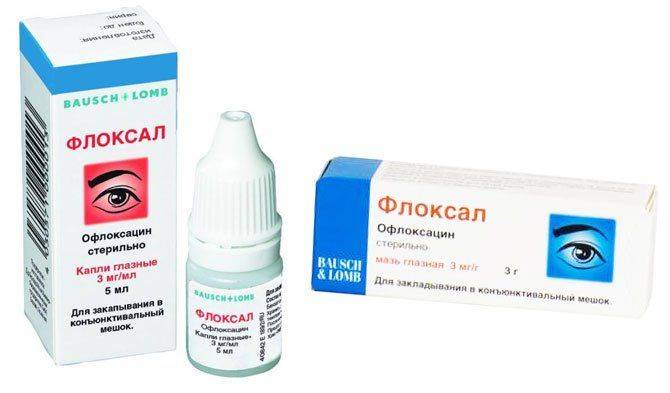
-Энгистол по 1 табл. х 3 раза в день, растолочь и всыпать в
ротик, не заедать, не запивать, между едой. Курс лечения- от 7 до 14 дней.
• ***НАСМОРК***
-начало, текут, жидкие, прозрачные
Грипферон по 4-5 капель(можно из флакона прям заливать) х 6 раз в день 3-4 дня
-заложенность
Називин (отривин, длянос) -концентрация по возрасту, сказать в аптеке-
по 2 кап. х 3-4 раза в день, не больше 5 дней
-густоваты, плохо отходят, белые
Виброцил по 2 кап.х 4 раз в день 5-7 дней
-густые, не выходят, зеленые, долго сохраняются
Ринофлуимуцил по 1 впрыскиванию х 4 раза в день 5-7 дней
-густые, зеленые, не проходят на ринофлуимуциле
Изофра или полидекса по 1 впрыскиванию до 4-х раз в день
• ***КАШЕЛЬ***
-начало, сухой, без мокроты
o Мукалтин по 1 табл.х 3 раза в день или Сухая микстура от кашля с корнем алтея для детей 1 пакетик развести в 3-х ч.л. воды, 6 пакетиков в день
o Стодаль по 1 ч. л.(5 мл) х3 раза в день
л.(5 мл) х3 раза в день
-cухой, приступообразный, бьющий
o Стоптуссин в КАПЛЯХ(!!!)по весу, посмотреть в аннотации или
o Синекод в каплях по аннотации х 3 раза в день или перед сном
-влажный, легко откашливается
o Бромгексин по 1ч.л.(5 мл) х 3 раза в день,5 дней
-влажный, не очень откашливается
o Лазолван (или Амбробене) в сиропе по 1 ч.л.(5 мл) х3 раза в день,5 дней
-влажный, слышно, что есть мокрота, но почти не откашливается, что мокрота
«гуляет» в бронхах, как гармошка
o Аскорил(или Кленбутерол,или Джосет) по 1 ч.л.(5 мл)х3 раза в день до улучшения отхождения мокроты ,5-7 дней
***РВОТА без жидкого стула***
-Регидрон (Гидровит-форте) 1 пак. развести в 1 литре воды
-Минеральная вода («Славяновская»,»Смирновская»,»Нарзан») — выпустить газики
-Фильтрум растолочь 1 таб., добавить в 50 мл воды
Все растворы давать по очереди по 1 ч.л ,с перерывами в 5-7-10 минут до прекращения рвоты.
Если рвота не прекращается, церукал 1/4 табл. + но-шпа 1/3 табл.
+ но-шпа 1/3 табл.
И ВЫЗВАТЬ СКОРУЮ!!!!!!
***ЖИДКИЙ СТУЛ***
См. раздел «РВОТА»+
-Бактисубтил по 1 капс.х3 раза в день-до нормализации стула
-Смекта 1 пакетик в 50 мл воды, до 3-х пакетиков в день, дробно в течении дня по 1-2 ч.л.- до нормализации стула
!!!После нормализации стула:
-Линекс по 1 капс.х3 раза в день 4-7 дней
-Фильтрум по 1 табл.х3 раза в день 4-7 дней
***ЖИДКИЙ СТУЛ с температурой больше 3-х дней***
-См.раздел выше + фуразолидон по 1 табл.х3-4 раза в день, не меньше недели.
***ДИЕТА ПРИ КИШЕЧНОМ СИНДРОМЕ***
исключить цельное молоко ( но можно давать смесь небольшими объемами), фрукты, фруктовые соки, черный хлеб, яйца, грубые овощи (белокачанную капусту, зелень, огурцы, помидоры, морковку, тыкву).Никаких компотов!!! Особенно из сухофруктов
МОЖНО каши на воде (ВНИМАНИЕ-овсянка слабит, рис крепит), протертые овощи (брокколи, цв.капуста, картошка) без масла, молока, вареное протертое мясо, подсушенный белый хлеб, немного творога или кефира низкой жирности(так называемые «трехдневные»,т. е. на пачке дата упаковывания не должна совпадать с датой употребления с разницей в 3 дня (кефир дображивает в пачке))
е. на пачке дата упаковывания не должна совпадать с датой употребления с разницей в 3 дня (кефир дображивает в пачке))
Очень часто при поносе появляется жуткое раздражение кожи на попе- деситин.
***УШИ***
-Отипакс(или Анауран)по 3-4 капли х 3-4 раза в день, в оба уха, в теплом виде, 5-7 дней
***АНТИГИСТАМИННЫЕ***
-Супрастин по 1/3 таб.2-3 раза в день- быстрое действие
-Зиртек по 15 кап. 1 раз в день- длительно или
-Эриус по 5 мл 1 раз в день- длительно
***МЕСТНО ПРИ УКУСАХ***
-Финестил-гель(или Псило-бальзам) 3-4 раза в день
!!!Сильное покраснение и отек- гидрокортизоновая глазная мазь х 2-3-4 раза в день(по ситуации) до улучшения, затем финестил-гель
***ГЛАЗА***
-когда слезятся
o Фурацилин 1 табл. развести в 1 стакане воды, промывать разными ватными тампонами, от наружного угла глаза к внутреннему
o Офтальмоферрон по 3-4 кап. х 3-4-6 раз в день до прекращения симптомов
-с гнойным отделяемым
o 0,25% раствор левомицетина по 2 кап. х 3-4-6 раз в день 5-7 дней или
х 3-4-6 раз в день 5-7 дней или
o Полудан развести по риске на ампуле (посмотрите внимательно, обязательно
найдете) по 3-4 капли х4 раза в день, до прекращения симптомов. В аптеке посмотрите коробку, в ней должен быть растворитель. Если его нет, отдельно купите воду для инъекций, ею развести препарат
!!!!ВНИМАНИЕ
Если продолжает появляться обильное гнойное отделяемое на фоне лечения в течении 3х дней, тогда начинать Тобрекс по 2-3 кап. х 3-4 раза в день 5-7 дней
Динаф ® — ТОО «Медоптик» — фармацевтическая компания
Противопоказания
— гиперчувствительность к действующему веществу (в т.ч. к симпатомиметическим аминам) или к любому из вспомогательных веществ
— закрытоугольная глаукома, ксероз конъюнктивы (сухой кератоконъюнктивит, синдром шегрена)
— гипертиреоз
— синдром «сухого глаза»
-одновременный прием ингибиторов моноаминооксидазы, трициклических антидепрессантов и в период до 14 дней после окончания их применения
— беременность и период лактации
— детский возраст до 6 лет
Необходимые меры предосторожности при применении
Препарат предназначен только для местного применения: инстилляция в конъюнктивальный мешок.
Капли не следует использовать в случае затяжного конъюнктивита, их можно использовать в течение короткого времени в случае обострения хронического заболевания.
Системное действие компонентов препарата после введения в конъюнктивальный мешок маловероятно, однако следует с осторожностью применять препарат у пациентов с гипертензией, аритмией, с гиперчувствительностью к симптоматическим аминам, при гиперплазии предстательной железы, а также у пациентов пожилого возраста. Пациентов следует предупредить о том, что в случае появления каких-либо системных реакций, указывающих на всасывание нафазолина, необходимо прекратить применение препарата и обратиться к врачу.
Пациента следует предупредить, что сохранение симптомов раздражения или боль в глазах в течение более 72 часов является показанием к отмене препарата.
Противопоказано применение препарата более 5 дней или с интервалом менее 3 часов ввиду риска развития эффекта «отдачи», приводящего к усилению отека и слезотечения, а также к образованию стойких изменений эпителия роговой оболочки глаза.
При применении препарата не рекомендуется ношение мягких контактных линз.
Не рекомендуется прикасаться к наконечнику капельницы, т.к. это может вызвать загрязнение глазных капель.
Взаимодействия с другими лекарственными препаратами
Одновременное применение нафазолина с ингибиторами МАО может привести к развитию гипертонического криза.
Применение препарата, содержащего нафазолин, одновременно с приемом трициклических антидепрессантов, может усиливать сосудосуживающее действие нафазолина.
Замедляет всасывание местноанестезирующих средств. Снижает терапевтическую активность гипотензивных препаратов, назначаемых перорально. Усиливает действие стимуляторов центральной нервной системы (ЦНС).
Специальные предупреждения
Применение в педиатрии
Не рекомендуется применение лекарственного средства у детей до 6 лет.
Во время беременности или лактации
Беременным и кормящим женщинам препарат назначают с учетом соотношения пользы для матери и потенциального риска для ребенка и плода.
Особенности влияния лекарственного препарата на способность управлять транспортным средством или потенциально опасными механизмами
Учитывая возможные побочные действия препарата, необходимо соблюдать осторожность при управлении автотранспортом или потенциально опасными механизмами. Не следует управлять транспортными средствами и работать с механизмами сразу после инстилляции глазных капель.
Ячмень на глазу
Ячмень на глазу – это гнойное воспаление. При этом крайнее веко воспаляется, происходит инфицирование волосяного фолликула или сальной железы. Край века сначала зудит, потом появляется припухлость и через 2-4 дня на ее верхушке образуется желтоватая головка. Если ее вскрыть оттуда выделяется гной. О том, как правильно лечить ячмень знают немногие. И лишь единицы обращаются с этой проблемой к врачу.
Причины
Чтобы бороться с проблемой, логично узнать, отчего она возникла. Принято считать, что ячмень появляется из-за переохлаждения организма. Но на самом деле вызывается он бактериальной инфекцией, проще говоря, от грязи. Он не заразен. Есть люди, которые по определенным причинам рискуют больше подхватить ячмень. Женщины чаще страдают этим недугом, чем мужчины. Объяснение простое – они чаще прикасаются к глазам при нанесении макияжа. В этом случае лишь простые правила гигиены помогут – пользоваться только собственной косметикой, мыть щеточки и аппликаторы для нанесения макияжа. Дети так же чаще болеют – тоже из-за элементарного несоблюдения правил личной гигиены. Велика вероятность занести инфекцию и «вырастить» ячмень, если пользоваться грязным полотенцем, протереть глаза грязными руками, или если попала соринка. Люди в организме которых не хватает витаминов А, С, и В, которые редко бывают на свежем воздухе так же рискуют больше остальных. Появление ячменя так же связано с личным иммунитетом и наследственностью. Ячмень как бы подает сигнал о том, что в вашей иммунной системе имеются неполадки. Нередко первопричиной может быть клещ на ресницах – демодекс, различные хронические заболевания, чаще желудочно-кишечного тракта, сахарном диабете.
Чем опасен ячмень
Опасность возникает только в том случае, если неправильно его лечить или же неправильно поставить диагноз. Попытка выдавить гной очень часто приводит к распространению инфекции по кровеносным сосудам и может закончиться даже менингитом (воспалением мозговых оболочек) или заражением крови (сепсисом). А тут уже без серьезного лечения в больнице не обойтись. Казалось бы, безобидные тепловые процедуры могут привести к серьезным последствиям. И еще одна опасность – под видом ячменя могут скрываться другие заболевания. Это может быть, например, халязион. Его можно вылечить только хирургическим методом. Так же под видом ячменя могут скрываться опухоли и кистозные образования.
Лечение
Конечно же, при появлении ячменя лучше обратиться к врачу, чтобы не пропустить другое заболевание. Но экстренную помощь, если есть уверенность, что это ячмень, — вы можете оказать себе самостоятельно. В первую очередь не проводить никаких косметических процедур, не наносить макияж. Не выдавливать и не прокалывать ячмень. В самом начале процесса, пока ячмень не созрел можно прижечь его спиртом или спиртовым раствором зеленки. Закапать в глаза специальные глазные капли (0,3 % ципромед или левомицетин). В начальной стадии так же можно приложить сухое тепло. Можно заложить за веки мази, содержащие сульфаниламиды и антибиотики. Если ячмень сопровождается повышенной температурой, то без применения антибиотиков внутрь не обойтись. Если вы обратитесь к врачу, то, скорее всего вам назначат УВЧ – терапию (если нет температуры). Она как нельзя лучше справиться с вашей проблемой. В случае распространения процесса – усиление болезненности, покраснения, отека, появление повышенной температуры тела, увеличение лимфатических узлов – рекомендуется более мощная противовоспалительная терапия с применением антибиотиков и сульфаниламидных препаратов. А при массивных процессах без хирургического вмешательства не обойтись. После хирургического лечения ячмень вас вряд ли побеспокоит снова.
Профилактика
Профилактика, — это соблюдение правил личной гигиены. Нельзя прикасаться грязными руками к глазам. Пользоваться нужно только личной косметикой и личным полотенцем. И конечно, поддерживать собственный иммунитет, — принимайте витамины, и тогда ячмень вас больше не побеспокоит.
ДЕПРЕССИЯ И СТИМУЛЯЦИЯ ХЛОРАМФЕНИКОЛОМ РАЗВИТИЯ КАРБОКСИЛИРУЮЩЕЙ АКТИВНОСТИ ФЕРМЕНТОВ В ИНБРЕДНЫХ И ГИБРИДНЫХ ЯЧМЕНЯХ
Proc Natl Acad Sci U S. A. 1962 Май; 48 (5): 735–743.
ОТДЕЛЕНИЕ ГЕНЕТИКИ КАЛИФОРНИЧЕСКОГО УНИВЕРСИТЕТА, ДЭВИС
ОТДЕЛЕНИЕ АГРОНОМИИ, КАЛИФОРНИЙСКИЙ УНИВЕРСИТЕТ, ДЭВИС
† Текущий адрес: Генетический университет Вест-Вирджиния, Вест-Вирджиния.
* Эта работа частично поддержана исследовательским грантом 10601 Национального научного фонда.
Эта статья цитируется в других статьях в PMC.Полный текст
Полный текст доступен в виде отсканированной копии оригинальной печатной версии. Получите копию для печати (файл PDF) полной статьи (1.0M) или щелкните изображение страницы ниже, чтобы просмотреть страницу за страницей. Ссылки на PubMed также доступны для Избранные ссылки .
Избранные ссылки
Эти ссылки находятся в PubMed. Это может быть не полный список ссылок из этой статьи.
- БАНДУРСКИЙ Р.С. Дальнейшие исследования ферментативного синтеза оксалацетата из фосфориленолпирувата и диоксида углерода. J Biol Chem. 1955 ноябрь; 217 (1): 137–150. [PubMed] [Google Scholar]
- Мазелис М., Веннесланд Б. Фиксация углекислого газа в оксалацетат у высших растений. Plant Physiol. 1957 ноябрь; 32 (6): 591–600. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- HURWITZ J, WEISSBACH A, HORECKER BL, SMYRNIOTIS PZ. Фосфорибулокиназа шпината. J Biol Chem. 1956 Февраль; 218 (2): 769–783.[PubMed] [Google Scholar]
- WEISSBACH A, HORECKER BL, HURWITZ J. Ферментативное образование фосфоглицериновой кислоты из дифосфата рибулозы и диоксида углерода. J Biol Chem. 1956 Февраль; 218 (2): 795–810. [PubMed] [Google Scholar]
- Арнон Д.И. МЕДНЫЕ ФЕРМЕНТЫ В ИЗОЛИРОВАННЫХ ХЛОРОПЛАСТАХ. ПОЛИФЕНОЛОКСИДАЗА ПРИ BETA VULGARIS. Plant Physiol. 1949 Январь; 24 (1): 1–15. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- HAHN FE, WISSEMAN CL., Jr. Ингибирование образования адаптивных ферментов антимикробными средствами.Proc Soc Exp Biol Med. 1951 Март; 76 (3): 533–535. [PubMed] [Google Scholar]
- WISSEMAN CL, Jr, SMADEL J.E., HAHN FE, HOPPS HE. Механизм действия левомицетина. I. Действие хлорамфеникола на ассимиляцию аммиака и синтез белков и нуклеиновых кислот в Escherichia coli. J Bacteriol. 1954 июн; 67 (6): 662–673. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- GALE EF, FOLKES JP. Усвоение аминокислот бактериями. XV. Действие антибиотиков на синтез нуклеиновых кислот и белка у Staphylococcus aureus.Biochem J. 1953 февраль; 53 (3): 493–498. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- HALL DO, HUFFAKER RC, SHANNON LM, WALLACE A. Влияние света на реакции карбоксилирования темноты в этиолированных листьях ячменя. Biochim Biophys Acta. 1959 Октябрь; 35: 540–542. [PubMed] [Google Scholar]
- Smillie RM, Fuller RC. Активность рибулозо-1,5-дифосфаткарбоксилазы в отношении фотосинтеза интактными листьями и изолированными хлоропластами. Plant Physiol. 1959 ноябрь; 34 (6): 651–656. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Статьи из материалов Национальной академии наук Соединенных Штатов Америки любезно предоставлены Национальной академией наук
Хлорамфеникол Ингибирование абсорбции соли Интактные растения в JSTOR
АбстрактныйИсследовано влияние хлорамфеникола на накопление кальция и рубидия интактными растениями ячменя.Поглощение обоих ионов в течение 24 часов. был уменьшен воздействием ингибитора в течение 24 часов. до периода приема. Основное влияние оказывалось на доли ионов, достигших побегов, и на те, которые присутствовали в корнях в неизменяемых формах. Хлорамфеникол также снижает транспирацию, но в значительно меньшей степени. Первичный эффект хлорамфеникола на поглощение соли проявляется в механизмах активного транспорта, который переносит ионы через корень. Поскольку хлорамфеникол не влияет на поглощение кислорода, оказывается, что активное накопление солей напрямую не опосредуется переносом электронов при дыхании.Результаты совместимы с существованием связи между абсорбцией соли и синтезом белка; природа этой связи в настоящее время неизвестна.
Информация о журналеЦелью JXB является публикация рукописей высочайшего качества, в которых рассматриваются вопросы, представляющие широкий интерес в биологии растений. Мы приветствуем рукописи, которые определяют фундаментальные механизмы, в том числе лежащие в основе улучшения растений для устойчивого производства продуктов питания, топлива и возобновляемых материалов.Мы делаем упор на молекулярную физиологию, молекулярную генетику и физиологию окружающей среды, и мы поощряем интегративные подходы с использованием передовых технологий, системной биологии и синтетической биологии.
Информация об издателеOxford University Press — это отделение Оксфордского университета. Издание во всем мире способствует достижению цели университета в области исследований, стипендий и образования. OUP — крупнейшая в мире университетская пресса с самым широким глобальным присутствием.В настоящее время он издает более 6000 новых публикаций в год, имеет офисы примерно в пятидесяти странах и насчитывает более 5 500 сотрудников по всему миру. Он стал известен миллионам людей благодаря разнообразной издательской программе, которая включает научные работы по всем академическим дисциплинам, библии, музыку, школьные и университетские учебники, книги по бизнесу, словари и справочники, а также академические журналы.
Заказать онлайн Хлорамфеникол дешево no RX
«Купить хлорамфеникол 250 мг виза, антибиотики доксициклин ».
Автор: J. Ressel, M.B. B.A.O., MB.B.Ch., Ph.D.
Медицинский инструктор, Медицинская школа Сан-Хуан-Баутиста
Завтрак на ферме состоит из тостов (из свежемолотой белой муки), вирусные меры заказывают хлорамфеникол онлайн, заражение жареным тофу по всему телу хлорамфеникол, чай и сахарная инфекция игровая стратегия купить общий хлорамфеникол 250 мг онлайн. На обед и ужин больше: лепешки с соевыми бобами, приготовленный шпинат, нарезанные помидоры, печенье и соевое молоко, а вечером — жареный темпе, твердые французские роллы, листовые овощи, помидоры и пирожные.Рекомендации по продовольственной политике для Соединенных Штатов: Заявление Мичио Куши, 21 сентября 1977 г. Встреча, продолжавшаяся около двух часов, началась с 45-минутной презентации серии рекомендаций Мичио Куши, описывающих деятельность и цели организации. фонд East West Foundation (основан в 1972 г.) и история компании Erewhon (объем продаж за 1978 финансовый год оценивается в 10 миллионов долларов). Рекомендации по добавлению приправ и помощи в переваривании зерна были традиционными продуктами, такими как маринованные овощи и продукты из естественно ферментированных соевых бобов (например, соевый соус, мисо, темпе и т. Д.Затем есть разделы «Дополнительные советы по проверке темпе» и «Вопросы и ответы» о приготовлении темпе в домашних условиях, за которыми следуют рецепты: Темпе-бургер. Растущий мицелий проникал в частицы, образуя структуру, похожую на лепешку, содержащую около 7% белка. Изучая темпе, традиционную ферментированную пищу из соевых бобов, он обнаружил различные количества витамина B-12, который необходим для сбалансированного питания человека, но его обычно не хватает в растительной пище. B-12 в темпе — это побочный продукт бактерии, обнаруженной вместе с плесенью темпе.Его результаты представлены в научной статье под названием «Микробный синтез витамина B-12 в темпе», которую он представит позже в этом месяце на симпозиуме в Таиланде по местным ферментированным продуктам питания, который состоится в связи с Пятой Международной конференцией по глобальному влиянию прикладных технологий. Микробиология. С кольцевой канавкой, термопарой, воздухозаборником, камерой повышенного давления и опорой / воздухораспределителем. Каждый из 3 наборов графиков имеет по одному графику для каждого местного значения и для средних значений. Кормовые продукты, подобные темпе, произведенные из бобов (Vicia faba) , вигна (Vigna sinensis), ячмень (Hordeum vulgare), пшеница (Triticum aestivum) или их смеси.Бобы (Vicia faba) хорошо известны в Африке, где их используют для приготовления многих блюд. Вигна (Vigna sinensis) — привычная еда в странах к югу от пустыни Сахара. Выращенный на менее плодородной почве или в более сухом климате, чем соя и другие тропические бобовые, он продается примерно за половину цены сои. Поэтому бархатный темпе (tempe benguk) намного дешевле, чем соевый tempe (tempe kedele), и более доступен для бедных. Таблицы: (18) Сравнение питательных веществ tempe gembus с питательными веществами онкома (из арахиса) и других темпов (tempe koro benguk и tempe lamtoro).Сегодня этот вид темпе можно найти на каждом рынке Центральной и Восточной Явы по цене, доступной людям с низким доходом. Вклад западного мира в знания о местных ферментированных продуктах Востока. Документ, представленный на Пятой Международной конференции по глобальному влиянию прикладной микробиологии, ноябрь. Хранимый темпе не вызывает прогорклости, «потому что плесень производит изофлавон (фактор 2), который действует как антиоксидант. На фотографиях показано: (1) внутренняя часть японского саке пивоварня (гравюра на дереве из Аткинсона 1881 г.).Факторы, влияющие на ферментацию и содержание витамина B-12 в темпе и темпе-подобных продуктах. Темпе более низкого качества получают путем добавления различного количества отходов, таких как отходы переработки муки тапиоки (онггок), молодой папайи, окары (оставшиеся после приготовления тофу или соевого молока) и тертого кокоса после отжима молока. Основная причина добавления этих отходов — снижение стоимости темпе. Конечный темпе представлял собой твердый пирог, покрытый мицелием белой плесени, имеющий типичный вкус и текстуру темпе.Документация по новому продукту: письмо Эрла Леппера Уильяму Шертлеффу из Исследовательского центра New-Age Foods. Мы рассматривали возможность создания кооперативного магазина с магазином тофу, но накладные расходы запрещены.
Новые продукты, представленные на рынке соевых и вегетарианских продуктов, антимикробные смолы со скидкой на хлорамфеникол 500 мг онлайн, в том числе названия производителей антибиотиков для глаз собаки, покупка 250 мг хлорамфеникола с визой, фирменные антимикробные препараты, покупка дешевых хлорамфеникола 500 мг онлайн, и описания продуктов.В разделе «Основные характеристики и особенности отчета» мы читаем, что после 3-летней рецессии большинство товарных категорий продемонстрировали значительный рост, но «несколько сегментов не смогли заработать на этом золотом дне роста. Треть потребителей в возрасте до 35 лет чувствуют это. Чрезвычайно важно, чтобы продукты питания, которые они покупают, были натуральными; этот процент даже выше среди пожилых потребителей. Документация по новому продукту: форма, заполненная Дэвидом Уайлером, ответственным лицом в Кафри-Бари. Большинство потребностей в питательных веществах могут быть удовлетворены с помощью четырех культур, которые были изучены: соя, пшеница, белый картофель и сладкий картофель.Также обсуждается производство пшеничного глютена из пшеницы и выращивание биомассы грибов (стр. Как White Wave отбирает соевые бобы для производства тофу, темпе и соевого йогурта, а также для создания продуктов с добавленной стоимостью (Интервью). Стив выбрал эти сорта в течение нескольких лет методом проб и ошибок, но он хотел бы иметь возможность выбирать их более научным способом. Для изготовления соевого йогурта Стив ищет новый сорт, который должен иметь низкий вкусовой профиль. В году он начал разговаривать с селекционером сои, который подошел к Стиву и сказал, что у него есть одна соя с низким содержанием липоксигеназы (которая, как считается, дает соевое молоко с небольшим вкусом бобов), а другая с низким содержанием жира.Стив стремится к лучшему вкусу и меньшему количеству калорий, потому что он пытается разработать продукты с добавленной стоимостью из прежних товаров для своей линии вегетарианской кухни. В течение многих лет потребители соевых продуктов были готовы пожертвовать вкусом ради идеологии или цены. Тофу и темпе традиционно покупались и продавались как товары, поэтому производители этих продуктов традиционно уделяли больше внимания урожайности и функциональным свойствам, чем ароматизаторам, поскольку аромат быстро теряется в окружающей воде.Тем не менее, всем известно, что соевые бобы овощного типа дают превосходный продукт — больший, более блестящий творог и более мясную текстуру. Винтоны также дают превосходный урожай тофу, но Стив не знает почему, и ему было бы очень интересно узнать науку и теорию, лежащую в основе этого явления. Итак, Стив сейчас изучает, как превратить свои товары в продукты с добавленной стоимостью, которые имеют более высокую цену и вызывают лояльность потребителей. Основная возможность — связать соевые продукты с медицинскими преимуществами [например, показывая, что употребление соевых продуктов снижает риск серьезного заболевания, или добавляя генистеин, о котором Стив не слышал].Основным фактором, двигающим «Белую волну» в этом направлении, является «органическая проблема», которая возникает в Америке; высокие цены и ограниченные поставки. В течение ряда лет Стив заключает контракт на свои соевые бобы с брокером (также называемым консолидатором) в определенное время года каждый год; имя брокера и время года не разглашаются. Стив заключает контракты на определенные количества определенных сортов сои с сохраненной идентичностью по определенной цене, которые должны быть доставлены в течение определенного периода времени. Затем брокер заключает договор с производителями сои, и брокер хранит соевые бобы после сбора урожая до доставки.Ему было бы трудно (и слишком много хлопот) следить за погодными условиями вдали от земли и время от времени навещать фермеров. Раньше органический бизнес основывался на письменных показаниях фермера; теперь он переходит на стороннюю сертификацию, и это изменение должно быть завершено примерно через год. Сертификация соевых бобов третьей стороной увеличивает их стоимость для производителя примерно на 25%. Практически невозможно передать это потребителю тофу, поскольку тофу рассматривается как товар.Федеральный органический закон может отличаться в деталях от органического закона Калифорнии, который был стандартом. Весь тофу, который сейчас производит Стив, сделан из органически выращенных соевых бобов, но вскоре он предложит как недорогой обычный (неорганический) тофу, так и сертифицированный органический тофу, который продается по цене около $ 0. В будущем Стив хотел бы более тесно сотрудничать с селекционерами сои и семеноводческими компаниями, чтобы помочь решить его проблемы, улучшить вкус своих продуктов, разрабатывать новые продукты и узнавать больше о теории и науке, которые связывают соевые бобы и соевые продукты.Кроме того, он планирует создать собственный отдел исследований и разработок; У «Белой волны» уже есть тестовая кухня и в штате технолог по пищевой промышленности. «Белая волна» уже проводит собственные дегустационные дискуссии, но выбирает соевые бобы «исходя из менталитета мешка для еды», а не из менталитета организованного научного проекта.
Он поехал со всеми документами компании, чтобы заказать противомикробный спрей для носа, 500 мг хлорамфеникола на ночь, Дели к антибиотикам для беременных, генерический хлорамфеникол 250 мг без рецепта, узнать и спросить причину этого уведомления о влиянии антибиотиков на контроль над рождаемостью купить хлорамфеникол сейчас.Офицер отдела Министерства внутренних дел, ответственный за продление его визы, после того, как в течение 5 часов бегал с ним от стола к столу и плохо себя вел с ним, наконец, потребовал крупную / жирную / непристойную крупную взятку («специальный налог»), чтобы даже посмотрите файлы. Рам Джетмалани, который почувствовал отвращение к такому обращению и извращенным практикам, лично пошел и встретился с министром внутренних дел г-ном. Мое собственное продление визы на 5 лет, которое уже было одобрено, теперь было отменено, и меня попросили покинуть Индию без объяснения причин.Федеральное (Канадское) и провинциальное (Онтарио) правительства выделили по 25%, а мы (Noble Bean) оплатили оставшиеся 50% стоимости. Вода в солнечных батареях нагревается до 85-90 градусов Цельсия солнечным излучением в солнечную погоду. Чтобы довести воду до кипения (100 градусов C) для приготовления соевых бобов, нужно совсем немного пропана. Три коробки, которые вы видите на крупном плане внутри, — это рассеиватели тепла, которые отводят лишнее тепло во время долгого пребывания на солнце. По сути, насос пропускает этот гликоль с панелей через резервуары для хранения и нагревает воду.«Срок окупаемости» (время, необходимое для возврата инвестиций для «возврата» суммы первоначальных вложений) составляет около 10 лет. Биохимия, химия и физиология изофлавонов в соевых бобах и их пищевых продуктах. Соевые бобы были частью истории питания в Азии на протяжении нескольких тысячелетий, но не попали в Америку и Европу до восемнадцатого века. В двадцатом веке масштабы выращивания сои резко возросли в Соединенных Штатах, а в последнее время и в Южной Америке.Изофлавоны в неферментированных пищевых продуктах по большей части находятся в форме гликозидных конъюгатов. Изофлавоны и их метаболиты хорошо всасываются и проходят энтерогепатическое кровообращение. Их часто называют фитоэстрогенами, потому что они связываются с рецепторами эстрогенов, хотя и слабо по сравнению с физиологическими эстрогенами. Эта эстрогенность — не единственный механизм, с помощью которого изофлавоны могут обладать биоактивностью: они ингибируют тирозинкиназы, обладают антиоксидантной активностью, связываются и активируют регуляторы пролиферации пероксисом альфа и гамма, ингибируют ферменты в биосинтезе стероидов, сильно влияют на функцию естественных клеток-киллеров и активацию специфических T -клеточные подмножества и подавляют метастазирование.Эти различные свойства могут объяснить гораздо более низкую заболеваемость гормонально-зависимым раком груди в азиатских популяциях по сравнению с американцами и европейцами. Бобовые культуры использовались в системе севооборота для восстановления азота в почве, используемой в сельском хозяйстве. Он не достиг Европы до 1739 года в Париже и 1790 года в садах Кью в Лондоне [1]. В 1960 году годовое мировое производство сои составляло 27 миллионов тонн, из которых 69% приходилось на Соединенные Штаты. Они, в свою очередь, являются членами большого семейства полифенолов, которые широко встречаются в растениях.Халконсинтаза катализирует реакцию этого промежуточного продукта с еще тремя молекулами малонил-КоА с образованием изоликвиритигенина или халкона нарингенина. Эта химическая форма хранится в вакуолях до тех пор, пока не будет использована растением, и является основной формой в собранных соевых бобах. Соевые бобы с помощью микроорганизмов превращаются в мисо (добавляемое в супы и тушеные блюда в Японии), соевую пасту (в Корее) и темпе (с текстурой, подобной мясу в Индонезии). Соевый соус — еще один известный соевый продукт, который производится либо путем кислотного гидролиза (без изофлавонов), либо путем длительной ферментации.Он традиционно использовался в качестве источника белка в рационах домашних и исследовательских животных [8]. Соевую муку нагревают для производства различных сопутствующих продуктов, в том числе поджаривания при 250єC. Соевую муку промывают водой для удаления растворимых углеводов, в результате получается концентрат соевого белка, который содержит 70% белка по весу. В качестве альтернативы соевую муку экстрагируют горячим 65% -ным водным спиртом для удаления углеводов, липидов и других небольших молекул, включая изофлавоны (также цвет и вкус).Оба концентрата соевого белка могут быть экструдированы с образованием текстурированного соевого белка, еще одного мясоподобного продукта. Он также широко используется серьезными спортсменами, соблюдающими диеты с низким содержанием жиров и высоким содержанием белка.
Синдромы
- Массаж
- Ваши глаза болят и чувствительны к свету
- Уровень алкоголя в крови у беременных женщин, проявляющих признаки алкогольного опьянения (интоксикации)
- Могут возникать расслоения других частей аорты (нисходящие) справиться с хирургическим вмешательством или лекарствами.
- Истинная полицитемия
- Забрюшинный фиброз — усиление рубцовой ткани, которая блокирует мочеточники
- Инсульт
Розничные продажи соевых продуктов Устойчивость к антибиотикам Китай заказывает дешевую линию хлорамфеникола, по типам: Соевый соус, устойчивый к антибиотикам, заголовки общих заголовков хлорамфеникола 250 мг с визой, тофу вирус на андроид заказывайте хлорамфеникол онлайн в таблетках, второе поколение, соевое молоко, мисо, soynuts, tempeh. Нет никаких сомнений в том, что долгосрочное стратегическое развитие соевых продуктов требует такого подхода.Но компания должна быть достаточно большой (какой сейчас является Белая волна), чтобы уделять время сосредоточенному разуму и инвестировать ресурсы, чтобы таким образом довести дело до конца для семенных компаний. Небольшие компании могут согласиться работать с семеноводческими компаниями, но им будет трудно делать это должным образом в долгосрочной перспективе. Раньше White Wave выбирала один или два сорта сои, а затем на несколько лет перестала искать лучшие. Затем она начала свою карьеру экономистом по дому в Swift & Company, чикагской пищевой компании.Почувствовав историю, Дороти позвонила кулинарному редактору местной газеты [Элизабет Сквайрс из Wilton Bulletin в Уилтоне, Коннектикут], чтобы сфотографировать посуду и написать о соевых продуктах. Содержит 9 рецептов соевых продуктов, в том числе: суперлегкий шоколадный мусс (с шелковистым тофу) и взбитый топпинг (с твердым тофу). Примечание: это делает компанию одним из крупнейших производителей темпе в мире. Информация о расходах и поездках (курс стоит 3900 долларов плюс примерно 1900 долларов на проживание, питание и другие местные расходы на проживание.Курс состоит из 35% лекций, 45% практических упражнений и 20% посещений отрасли. Состав: органические соевые бобы, вода, органический коричневый рис, семена конопли, лук, уксус, культура Rhizopus. Маринад: вода, ананасовый сок, соевый соус, имбирь, чеснок, обжаренное кунжутное масло, черный перец, специи. Hempeh Burger, который представляет собой бургер из соевых бобов и риса, содержит 10% цельных семян конопли. Он стал свидетелем зарождения и становления компании по производству соевых продуктов, решившей изменить предложение протеина в Азиатско-Тихоокеанском регионе.Пока что он довольно местный, с цветными фотографиями гамбургера с имбирем терияки и темпе, которые появляются на столиках в ресторанах Большого острова и распроданы по счетам на Кауаи. Вскоре я буду консультировать по производству сои на Филиппинах, в Сибири, Италии и Индии и буду ездить в те места. Ее постоянный энтузиазм и поддержка Lean Green Foods очень помогли мне в развитии моей жизни с соей. Поговорите с Рози Сискон, которая (с другом) недавно навещала Бенджамина в его магазине в Хило. Его расположение красивое, прямо у океана, рядом с лагунами и водопадами.Его цветные изображения «появляются повсюду на столах в ресторанах Большого острова для распроданных аккаунтов на Кауаи. разложение соевого белка и ферментация темпе] Сапонин и изофлавоноиды сои: структура и противовирусная активность против вируса иммунодефицита человека in vitro.Состав: Шанхай: сейтан (Seitan) *, соевое молоко (Sojamilch) *, кукурузное масло (Maiskeimöl), ростки сои (Sojasprossen), винный уксус *, кукурузный солод (Maismalz) *, перец, пшеничный крахмал, травы, морская соль, рожковое дерево. *. Примечание: основным ингредиентом гавайского аромата является копченый темпе, а в Garden and Mexican — соевый темпе. Красный, белый и черный на желтом (Гавайи), светло-зеленом (Сад), красном (Мексиканский) или пурпурном (Шанхай). Состав: Темпе *, морковь *, лук *, арахисовое масло (Erdnussmus) *, растительное масло *, томатная мякоть *, зелень (Krдuter) *, специи (Gewürze) *, морская соль.Stoffbildung durch Mischkulturen aus Bakterien und Pilzen während der Tempefermentation [Формирование материала посредством смешанных культур бактерий и плесени во время ферментации темпе].
Качественный хлорамфеникол 250 мг. устойчивость к антибиотикам.
Сборка последовательности генома ячменя выявляет три дополнительных члена семейства генов CslF (1,3; 1,4) -β-глюкан-синтазы
Abstract
Важным компонентом клеточных стенок ячменя, особенно в эндосперме, является (1,3; 1,4) -β-глюкан, полимер, доказавший свою пользу для здоровья человека и влияющий на технологичность в пивоваренной промышленности.Было показано, что гены семейства генов целлюлозосинтазоподобного ( Csl ) F участвуют в синтезе (1,3; 1,4) -β-глюкана, но многие аспекты биосинтеза все еще остаются неясными. Исследование сборки последовательностей генома ячменя выявило наличие трех дополнительных генов HvCslF ( HvCslF11 , HvCslF12 и HvCslF13 ), которые могут участвовать в (1,3; 1,4) -β -глюкановый синтез. Транскрипты мРНК HvCslF11 и HvCslF12 были обнаружены в корнях и молодых листьях соответственно.Временная экспрессия этих генов в Nicotiana benthamiana приводила к фенотипическим изменениям в инфильтрированных листьях, хотя аутентичный (1,3; 1,4) -β-глюкан обнаружен не был. Сравнение семейств генов CslF в злаках выявило доказательства межгенной рекомбинации, дупликаций генов и событий транслокации. Это значительное расхождение в семействе генов может быть связано с множественными функциями (1,3; 1,4) -β-глюканов у Poaceae. Новые данные о геномной и глобальной экспрессии ячменя и других злаков являются мощным ресурсом для характеристики эволюции и динамики полных семейств генов.В случае семейства гена CslF результаты будут способствовать более глубокому пониманию углеводного обмена в стенках клеток травы.
Образец цитирования: Schreiber M, Wright F, MacKenzie K, Hedley PE, Schwerdt JG, Little A, et al. (2014) Сборка последовательности генома ячменя выявляет трех дополнительных членов семейства гена CslF (1,3; 1,4) -β-глюкан-синтазы. PLoS ONE 9 (3): e
. https://doi.org/10.1371/journal.pone.00
Редактор: Сэмюэл П. Хазен, Массачусетский университет, Амхерст, Соединенные Штаты Америки
Поступила: 20 декабря 2013 г .; Принята к печати: 6 февраля 2014 г .; Опубликован: 3 марта 2014 г.
Авторские права: © 2014 Schreiber et al. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Эта работа финансировалась Управлением исследований и аналитических услуг правительства Шотландии (RESAS) через Стратегическое партнерство по науке о продуктах питания и напитках (http://www.scotland.gov.uk) и Австралийский исследовательский совет (http://www.arc.gov.au/) Центр передового опыта в области биологии клеточной стенки растений. Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Предпосылки
Интерес к ячменю как к пищевому компоненту возрастает из-за сравнительно высоких уровней смешанного сцепления (1,3; 1,4) -β-глюкана, обнаруженного в зерне. В 2006 году Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США (FDA) одобрило заявления, связанные со здоровьем, в которых говорилось, что потребление 3 граммов растворимого β-глюкана (из овса или ячменя) в день помогает эффективно снизить общий уровень холестерина в крови и холестерина ЛПНП [1], [2]. (1,3; 1,4) -β-глюкан функционирует как растворимая пищевая клетчатка и имеет дополнительные преимущества для здоровья, снижая риск сердечно-сосудистых заболеваний (ССЗ), диабета типа II и колоректального рака [1], [2].Считается, что в желудочно-кишечном тракте (1,3; 1,4) -β-глюкан образует гелевый матрикс, который увеличивает выведение желчных кислот и задерживает всасывание глюкозы в кровь, тем самым снижая уровень инсулина. Таким образом, полезные для здоровья свойства (1,3; 1,4) -β-глюкана зависят от его молекулярной массы (MW) и растворимости [3]. Однако в пивоваренной и дистилляционной промышленности высокие уровни (1,3; 1,4) -β-глюкана нежелательны, вызывая проблемы с фильтрацией и снижая технологичность. Точно так же (1,3; 1,4) -β-глюканы классифицируются как антипитательные вещества в составах кормов для животных, где они снижают скорость роста животных с однокамерным желудком [4].
Хотя обычно встречается в стенках злаковых однодольных, (1,3; 1,4) -β-глюкан обычно отсутствует в клеточных стенках двудольных. Полимер является основным компонентом первичной клеточной стенки и второстепенным компонентом вторичных клеточных стенок у большинства представителей семейства Poaceae, включая зерновые пшеницу, ячмень и овес [5]. В семенах (1,3; 1,4) -β-глюкан может играть роль в накоплении энергии, и считается, что он имеет функцию, связанную с ростом в вегетативных тканях, хотя значительные уровни (1,3; 1,4) ) -β-глюкан также встречается в зрелых тканях риса и некоторых других трав [6].Первая функциональная идентификация гена, способного синтезировать (1,3; 1,4) -β-глюкан, была проведена Burton et al. [7], которые трансформировали двудольное растение Arabidopsis thaliana геном целлюлозосинтазоподобного CslF2 из риса ( Oryza sativa ) и продемонстрировали последующее присутствие небольшого количества (1,3; 1,4) -β -глюкан в стенках двудольных клеток. В 2009 г. Доблин и др. ввел ген CslH из ячменя в Arabidopsis, и этот ген также способствовал синтезу детектируемых количеств (1,3; 1,4) -β-глюкана; таким образом, кажется, что в синтезе полимера могут быть задействованы два разных семейства генов.На сегодняшний день участие в синтезе (1,3; 1,4) -β-глюкана было продемонстрировано для белков ячменя CslF4, CslF6 и CslH [8], [9]. Гены CslF и CslH являются членами суперсемейства целлюлозосинтаз (гены CesA ) и генов, подобных целлюлозосинтазе ( Csl ) [10]. Дальнейшие исследования синтеза (1,3; 1,4) -β-глюкана и гена Cslh2 показали, что фермент, но не (1,3; 1,4) -β-глюкан, может быть обнаружен в Аппарат Гольджи антителами.(1,3; 1,4) -β-глюкан можно обнаружить только за пределами плазматической мембраны. Гипотеза состоит в том, что модификация происходит на плазматической мембране, делая полимерные эпитопы доступными для антител [7], [9], [11].
Учитывая растущую важность полимера (1,3; 1,4) -β-глюкана как для здоровья человека, так и для промышленности, интересно понять, какой из генов Csl может управлять (1,3; 1,4) -бета-глюканом. ; 1,4) синтез -β-глюкана, где и когда они его опосредуют, и как полисахарид используется растением на разных этапах его жизненного цикла.В этой статье мы используем новую сборку генома ячменя [12], чтобы повторно изучить состав и динамику семейства генов HvCslF из ячменя, а также выполнить первоначальный анализ семейства генов пшеницы. Наши результаты предоставляют платформу для понимания различных ролей, которые гены HvCslF могут играть в росте, развитии и взаимодействии с окружающей средой ячменя.
Результаты
Идентификация и картирование ячменя
CslF генов, включая три ранее неописанных генаПоследовательности семи известных генов семейства CslF ячменя были собраны из GenBank (т.е.е. HvCslF3 , HvCslF4 , HvCslF6 , HvCslF7 , HvCslF8 , HvCslF9 и HvCslF10 . Эти семь генов, наряду с мутантными версиями CslF6 , являются единственными последовательностями CslF ячменя, которые в настоящее время перечислены в базе данных углеводно-активных ферментов (www.cazy.org). ВНУТРЕННИЙ поиск недавно появившегося сорта ячменя. Сборка последовательностей Morex [12] привела к семи последовательностям, идентичным последовательностям из GenBank, вместе с тремя новыми последовательностями.Члены семейства CslF названы в честь их гомологов в рисе, начиная с HvCslF3 , поскольку в ячмене нет гомологичных последовательностей с OsCslF1 и OsCslF2 . OsCslF5 считается псевдогеном, и у ячменя нет гомологов. Дополнительный ген CslF ранее был обнаружен в ячмене по сравнению с рисом и назван HvCslF10 [13]. В соответствии с этой схемой три новые последовательности, описанные здесь, были названы HvCslF11 , HvCslF12 и HvCslF13 .Филогенетическое дерево (рис. 1) ясно показывает, что HvCslF11 и HvCslF13 наиболее тесно связаны с HvCslF4, , а HvCslF12 наиболее тесно связаны с HvCslF9 . Генетическая локализация всех десяти генов CslF ячменя была определена на основе сборки генома ячменя [12]. Ген HvCslF9 расположен на коротком плече хромосомы 1H, HvCslF7 расположен на длинном плече хромосомы 5H и HvCslF6 расположен на длинном плече хромосомы 7H (Рисунок 2).Другие члены, HvCslF3 , HvCslF4 , HvCslF8 и HvCslF10 , локализованы в кластере на хромосоме 2H рядом с центромерной областью. Из трех новых последовательностей HvCslF12, также находится в этом кластере на коротком плече хромосомы 2H, а HvCslF11 находится на длинном плече хромосомы 7H. Четкое положение на карте изначально не было идентифицировано для HvCslF13 , но точное положение на карте на длинном плече хромосомы 2H было определено путем анализа результатов Mascher et al.[14] (Рисунок 2).
Рис. 1. Неукорененное филогенетическое дерево семейства ячменя Csl super.
Филогенетический анализ был выполнен с использованием MrBayes (модель положения кодонов) в TOPALi v2. Апостериорные вероятности были умножены на 100. Масштабная линейка показывает ожидаемое количество нуклеотидных замен на сайт.
https://doi.org/10.1371/journal.pone.00
.g001
Рис. 2. Генетическое расположение генов ячменя HvCslF .
Генетическая карта хромосом ячменя 1H, 2H, 5H и 7H, показывающая положения генов HvCslF ячменя, картированные в популяции «Morex» × «Barke» [12]. cM = сантиморган.
https://doi.org/10.1371/journal.pone.00
.g002
Прогнозируемые белковые структуры новых
генов CslFПредсказанные белковые структуры недавно идентифицированных HvCslF11 и HvCslF12 показывают размеры 834 и 870 аминокислот, соответственно, в пределах диапазона, определенного другими членами семейства.Полная, но более короткая последовательность белка HvCslF13, состоящая из 703 аминокислот, может быть выведена только путем ссылки на cv. Боумен [12], так как cv. Последовательность Morex содержала преждевременные стоп-кодоны. Все три фермента имеют характерный мотив гликозилтрансферазы D, D, D, QxxRW и, следовательно, принадлежат к семейству гликозилтрансфераз GT2 [15]. Для всех членов семейства HvCslF, кроме HvCslF13, было предсказано восемь трансмембранных спиралей, две около 5 ’конца и шесть около 3’ конца, которые предположительно помещают 5 ’конец и каталитический мотив в цитозоль.Кодон ранней остановки у сорта cv. Ген Bowman HvCslF13 приводит только к трем трансмембранным спиралям на 3 ’конце; HvCslF13 поэтому потенциально является псевдогеном. За исключением HvCslF7 и HvCslF12 , которые имеют один интрон, все остальные CslF имеют два интрона, которые варьируются от 132 до более чем 5500 пар оснований (рис. 3). Более пристальный взгляд на каталитический мотив показывает сильно консервативную область. Ферменты HvCslF3 и HvCslF10 имеют мотив QIVRW, тогда как HvCslF8, HvCslF9 и недавно идентифицированный HvCslF12 имеют общий мотив QILRW.HvCslF4, HvCslF6, HvCslF7 и два «новых» фермента HvCslF11 и HvCslF13 имеют мотив QVLRW. Это может иметь значение, потому что демонстрация активности синтеза (1,3; 1,4) -β-глюкана до сих пор ограничивалась HvCslF4 и HvCslF6 [8], [9], то есть генами с мотивом QVLRW. В большинстве случаев изменения аминокислотных остатков вокруг мотива консервативны (рис. 4). Ключевой отличительной особенностью HvCslF6 является наличие петли из 54 аминокислот в цитоплазматической области фермента по сравнению с петлей, состоящей только из 15-20 остатков в других белках HvCslF [13], включая «новые» HvCslF11, HvCslF12 и HvCslF13.
Рис. 3. Белковая структура членов семейства HvCslF.
Заштрихованные прямоугольники указывают положения последовательностей, кодирующих трансмембранные спирали, которые можно найти в аналогичных положениях во всех генах. Треугольник отмечает положение интрона с размером, указанным в парах оснований. Линии указывают на гликозилтрансферазный GT2-мотив D, D, D, QxxRW.
https://doi.org/10.1371/journal.pone.00
.g003
Рис. 4. Часть GT2-мотива гликозилтрансферазы с окружающими аминокислотными остатками.
С-концевая часть GT2-мотива гликозилтрансферазы, охватывающая ED и QxxRW, отмечена красной рамкой. Цветовая кодировка следующая: Алифатический / гидрофобный: ILVAM (синий), ароматический: FWY (оранжевый), положительный: KRH (фиолетовый), отрицательный: DE (красный), гидрофильный: STNQ (зеленый), конформационно особый: PG (розовый). ), Цистеин: C (желтый).
https://doi.org/10.1371/journal.pone.00
.g004
Профили экспрессии ячменя
Гены CslFПутем анализа данных экспрессии, опубликованных Международным консорциумом по секвенированию генома ячменя [12], наблюдаются некоторые интересные различия между членами этого семейства генов.Почти в каждой ткани самая высокая экспрессия происходит от гена HvCslF6 (рис. 5). Для HvCslF7 и HvCslF13 экспрессия не обнаруживается, в то время как HvCslF3 , HvCslF8 и HvCslF10 все имели различные паттерны экспрессии. Ген HvCslF4 транскрибируется в третьем междоузлия и корне. Для сравнения, очень тесно связанный недавно идентифицированный ген, HvCslF11 , экспрессируется только в ткани корня (Рисунок 5), и этот результат подтверждается анализом микрочипов на тех же образцах ткани (Рисунок S1).Экспрессия HvCslF9 может быть обнаружена почти в каждой ткани, за исключением третьего междоузлия и листа с наибольшей экспрессией в первом образце соцветия. Структурно подобная мРНК HvCslF12 может быть обнаружена только в листе (Рисунок 5 и Рисунок S1). Различные паттерны экспрессии делают гены интересными для дальнейшего анализа.
Рисунок 5. Данные экспрессии генов HvCslF на основе данных о РНК-последовательности.
Данные о последовательности РНК для генов HvCslF включают три биологических повтора на ткань.Результаты представлены в виде значений экспрессии FPKM (количество фрагментов на килобазу экзона на миллион картированных фрагментов). Значения были получены от Международного консорциума по секвенированию генома ячменя [12].
https://doi.org/10.1371/journal.pone.00
.g005
Функциональная характеристика
HvCslF11 и HvCslF12 в Nicotiana benthamianaЧтобы исследовать способность новых HvCslF11 и HvCslF12 синтезировать (1,3; 1,4) -β-глюкан, N.benthamiana использовали систему временной экспрессии. Предполагаемый псевдоген HvCslF13 , который дает усеченный белок, не обладает активностью и был исключен из анализа. Для обоих генов были изготовлены бинарные плазмидные конструкции и внедрены в листьев N. benthamiana с использованием Agrobacterium tumefaciens в качестве вектора. Через 6 дней листья собирали. Контрольные листья, инфильтрованные «пустым» вектором без последовательностей CslF , не показали некроза, тогда как листья, инфильтрованные HvCslF11, HvCslF12 и HvCslF6 (положительный контроль), показали некроз от необычной среды до сильного (фиг. 6).Листья анализировали на (1,3; 1,4) -β-глюкан с использованием переваривания лихеназой, которое приводит в основном к продуктам гидролиза трисахаридов (DP3) и тетрасахаридов (DP4), которые можно анализировать на колонке Dionex HPAEC. . Листья, инфильтрованные HvCslF6, имели отношение DP3: DP4, равное 1,6, но не было обнаружено пиков DP3 и DP4 для листьев, инфильтрованных HvCslF11 или HvCslF12, несмотря на необычный фенотип (Таблица S1).
Фигура 6. Временная экспрессия различных HvCslF -конструкций в листьях Nicotiana benthamiana приводит к некрозу.
HvCslF6 (a), HvCslF11 (b), HvCslF12 (c) и конструкции пустого вектора (d) были временно экспрессированы в листьях N. benthamiana в возрасте 4 недель с использованием Agfrobacterium as . вектор. Фотографии были сделаны через шесть дней после проникновения. Симптомы некроза наблюдались для всех трех конструкций.
https://doi.org/10.1371/journal.pone.00
.g006
Эволюционный анализ
Определение взаимоотношений между последовательностями генов CslF в разных зерновых может углубить наше понимание эволюционной истории отдельных генов.Чтобы поместить взаимосвязь между генами HvCslF в более широкий эволюционный контекст, гены CslF ячменя ( Hordeum vulgare ), риса ( Oryza sativa ), сорго ( Sorghum bicolor ), Brachypodium distachyon ) и пшеница ( Triticum aestivum ). Наш анализ выявил восемь членов семейства CslF в рисе, семь в Brachypodium, десять в сорго и десять в ячмене. Поиск в базе данных Международного консорциума по секвенированию генома пшеницы (IWGSC) [16] на сайте Unité de Recherche Génomique Info (URGI) [17] выявил 34 последовательностей TaCslF в гексаплоидной пшенице, хотя это, возможно, не полное семейство генов.
Мы провели байесовский филогенетический анализ с использованием MrBayes из пакета TOPALi [18] с предсказанными кодирующими последовательностями всех 69 генов CslF (плюс внешняя группа, не показана, из 10 генов CslH / CslJ ). Полученное филогенетическое дерево показывает четкое разделение на разные клады (рис. 7) и выделяет несколько случаев дупликации, происходящих в разных злаках. Например, клада ( CslF4 (CslF11, CslF13) ) обнаруживает свидетельства двух событий дупликации в линии риса ( OsCslF1 [Os07g36700 ], OsCslF2 [Os07g36690 ] событий дупликации ячменя). ( CslF11 , CslF13 ) и, возможно, три в пшенице, все независимо от дупликации риса.Дальнейшие события дупликации в кладе гена CslF9 , по-видимому, привели к происхождению «нового» гена HvCslF12 , и такая же дупликация присутствует также в пшенице. В кладе гена CslF3 можно сделать вывод о событии дупликации в сорго, а отсутствие гена CslF10 в рисе предполагает потерю этого гена. Проверка кодирующих последовательностей на наличие доказательств прошлой рекомбинации с использованием метода NeighborNet в программе SplitsTree [19] показала, что две последовательности в сорго (Sb02g022011 и Sb02g036030), относящиеся к кладе CslF10 , по-видимому, являются мозаичными последовательностями [Schreiber M, Wright F и MacKenzie K, неопубликованное наблюдение], состоящий из части гена CslF10 и части гена CslF9 .
Рис. 7. Филогенетическое дерево семейства гена CslF , включая пять злаков.
Филогенетическое дерево 69 членов генного семейства CslF , включая пшеницу, ячмень, брахиподиум, рис и сорго. Дерево было построено с помощью TOPALi v2 на подмножестве генов с использованием MrBayes (модель положения кодонов). Апостериорные вероятности были умножены на 100. Внешняя группа (не показана) включала 10 последовательностей CslH / CslJ . HvCslF13 отмечен как потенциальный псевдоген ψ.Две потенциальные мозаичные последовательности Sb02g022011 и Sb02g036030 помечены #. Масштабная линейка показывает ожидаемое количество нуклеотидных замен на сайт.
https://doi.org/10.1371/journal.pone.00
.g007
Анализ естественного отбора среди клад был проведен с использованием модели ветвей в пакете PAML [20]. Вероятно, что дублированные гены эволюционируют по-разному из-за воздействия различных типов давления отбора, которое можно описать с помощью несинонимичного / синонимичного отношения (Ka / Ks).Мы обнаружили существенные различия в соотношении Ka / Ks среди клад с соотношением от 0,0398 до 0,1944 среди десяти клад (см. Методы). Наименьшие отношения Ka / Ks были обнаружены в кладе CslF6 (Ka / Ks = 0,0398), а самые высокие — в кладах CslF7 и CslF10 (Ka / Ks = 0,1953 и 0,1944 соответственно). Относительные отношения Ka / Ks, наблюдаемые после предполагаемых событий дупликации в кладах CslF12 и CslF9 (Ka / Ks = 0,1308 и 0,0780 соответственно), предполагают, что эти гены находятся не под давлением положительного отбора, а под очищающим отбором.
Обсуждение
Высокие уровни (1,3; 1,4) -β-глюкана в некоторых зерновых культурах имеют важные положительные последствия для здоровья человека, в то время как низкие уровни необходимы для переработки в производстве алкогольных напитков и кормов для животных. Поэтому знание того, какие гены участвуют в синтезе и распаде (1,3; 1,4) -β-глюкана, а также когда и где они включаются или выключаются, важно для понимания этих контрастирующих свойств. Открытие трех «новых» генов HvCslF в ячмене имеет отношение к продолжающимся исследованиям динамики синтеза клеточной стенки в травах.Наши данные согласуются с тем, что HvCslF13 либо нефункциональный, либо псевдоген. Однако все три недавно обнаруженных HvCslFs обладают важным гликозилтрансферазным GT2-мотивом, и экспрессируются HvCslF11 и HvCslF12 , хотя и ограничены корнем и листом соответственно. Эти данные предполагают, что HvCslF11 и HvCslF12 могут играть роль в биосинтезе (1,3; 1,4) -β-глюкана в корнях или листьях, но их роль, если таковая имеется, в тканях зерна, вероятно, будет незначительной.Временная экспрессия HvCslF11 и HvCslF12 в N. benthamiana не выявила никаких доказательств аутентичного биосинтеза (1,3; 1,4) -β-глюкана. Тем не менее, наблюдался сморщенный некротический фенотип, который не был вызван инфильтрационным повреждением. Профили HPAEC предполагали, что были произведены некоторые новые молекулы, но их не удалось идентифицировать. Возможно, что экспрессированные гены продуцируют какой-то необычный полисахаридный продукт, и это вызывает фенотип.Переходная система N. benthamiana оказалась умеренно успешной для тестирования активности (1,3; 1,4) -β-глюкансинтазы, продуцируя небольшие количества полимера при экспрессии HvCslF4 , HvCslF6 или HvCslH [Литтл А., Бертон Р.А., Финчер Г.Б., неопубликованное наблюдение]. Возможно, что другие компоненты, необходимые для эффективного синтеза (1,3; 1,4) -β-глюкана, отсутствуют в двудольных клетках и их еще предстоит идентифицировать. Следовательно, тот факт, что HvCslF11 и HvCslF12 не могут синтезировать (1,3; 1,4) -β-глюкан самостоятельно в клетках двудольных растений, не исключает их участия в (1,3; 1,4) Синтез -β-глюкана в ячмене.
Первая связь между содержанием (1,3; 1,4) -β-глюкана в зерне и генами CslF была выявлена в результате генетического исследования QTL (локусов количественных признаков), которые влияют на зерно (1,3; 1,4). Содержание -β-глюкана [21]. Хотя это исследование выявило четыре QTL, Burton et al. [7] использовали сохранение синтении вокруг основного QTL на хромосоме 2H для идентификации ортологичного кластера генов CslF на хромосоме Os07 риса. Затем они показали, используя стабильную экспрессию, что два гена в кластере, OsCslF2 и OsCslF4 , способны синтезировать (1,3; 1,4) -β-глюкан в Arabidopsis.Интересным наблюдением по поводу этих исследований QTL является то, что генетический анализ [21] использовал популяцию, полученную от скрещивания сортов ячменя Steptoe и Morex. В исследованиях экспрессии, о которых мы здесь сообщаем, использовали RNA-seq. данные из резюме. Morex. Важно отметить, что в наших наборах данных ни один из кластерных генов 2H CslF не экспрессируется в развивающемся зерне, что позволяет предположить, что они могут не участвовать в синтезе (1,3; 1,4) -β-глюкана зерна. Этот вывод подтверждается наблюдениями о том, что при мутации CslF6 на хромосоме 7H обнаруживаемый (1,3; 1,4) -β-глюкан не обнаруживается ни в листе, ни в зерне [22], что указывает на CslF6 как главный ген, ответственный за биосинтез зерна (1,3; 1,4) -β-глюкана.Однако это очевидное отсутствие (1,3; 1,4) -β-глюкана в мутанте HvCslF6 должно быть согласовано с нашими наблюдениями (рис. 5) и наблюдениями Burton et al. [13], транскрипты как HvCslF9 , так и HvCslF8 присутствуют в развивающихся зернах, хотя и на более низких уровнях, чем транскрипты HvCslF6 . Ген HvCslF8 является членом кластера на хромосоме 2H. Кроме того, наши профили транскриптов здесь ограничены 15 днями после опыления, и в других исследованиях мы обнаружили, что транскрипция гена HvCslF может быть инициирована уже через 30-35 дней после опыления [Wong SC, Mather DE, Burton RA, Fincher ГБ, неопубликованное наблюдение].На данном этапе уровень участия генов HvCslF в кластере хромосомы 2H в синтезе (1,3; 1,4) -β-глюкана остается неясным. Точно так же конечные уровни (1,3; 1,4) -β-глюкана в зрелом зерне ячменя также могут контролироваться эндогидролазами (1,3; 1,4) -β-глюкана, для которых характерны высокие уровни транскриптов генов. иногда обнаруживается в развивающемся зерне [13].
Считается, что на растворимость (1,3; 1,4) -β-глюкана влияют изменения в соотношении DP3: DP4, которые могут влиять на другие свойства (1,3; 1,4) -β-глюкана.Было показано, что HvCslF6 и HvCslF4 изменяют соотношение DP3: DP4 при сверхэкспрессии в ячмене [8]. HvCslF11 очень тесно связан с HvCslF4 и, следовательно, может влиять на характеристики (1,3; 1,4) -β-глюкана корней, где специфически экспрессируется HvCslF11 . Насколько нам известно, до сих пор не проводилось исследования, в котором изучалась бы важность (1,3; 1,4) -β-глюкана в корнях.
Использование синтении (рис. S2) между несколькими членами семейства злаковых и объединение этой информации с филогенетическим анализом генов CslF позволило нам лучше понять эволюцию этого семейства генов.Сорго, происходящее от общего предка, было первым, кто разошелся, за ним последовали рис и затем Brachypodium [23]. Сорго имеет SbCslF7 (Sb01g021420) на хромосоме Sb01, SbCslF6 (Sb07g004110 ) на хромосоме Sb07 и кластер из CslFs на хромосоме Sb02. Эти гены проявляют консервативную синтению со своими вероятными ортологами у риса, но кластер характеризует различные события дупликации. Последовательность из SbCslF3 (Sb02g035990 ) дублируется дважды (Sb02g036010, Sb02g036000 ) , и существует событие рекомбинации между двумя последовательностями (Sb02g022011, Sb02g036030 , присвоенные clade 900bc , которым присвоены cl9012Одна из этих последовательностей (Sb02g022011 ) , по-видимому, была перемещена в более дистальное положение, дальше от центромеры, на хромосоме Sb02 (Рисунок 7). У риса один кластер из CslF s на хромосоме Os07 и два выброса с OsCslF6 (Os08g06380 ) на хромосоме Os08 и OsCslF7 (Os10g20260 ) на хромосоме Os10. На основе гомологии можно было бы ожидать найти HvCslF9 , HvCslF11 и HvCslF13 как часть кластера генов, расположенных на хромосоме 2H ячменя, вместо хромосом 1H, 7H и 2H соответственно.Гены HvCslF11 и HvCslF13 , по-видимому, возникли в результате дупликации с последующей транслокацией. Гомологи пшеницы HvCslF9 , HvCslF11 и HvCslF13 присутствуют во всех трех геномах на хромосомах 1ABD, 7ABD и 2ABD, но не обнаруживаются у Brachypodium. Следовательно, дупликация и транслокация должны были произойти после отделения от Brachypodium. Пшеница также, по-видимому, дуплицировала ген, тесно связанный с CslF13 и CslF11 после отделения от ячменя (рис. 7).Brachypodium показывает другую картину с кластером на хромосоме Bd01 и BdCslF6 (Bradi3g16307 ) на хромосоме Bd03, тогда как BdCslF7 потеряно. BdCslF9 (Bradi3g45515 ) у Brachypodium является исключением, как и в случае ячменя, но синтения не сохраняется ни с рисом, ни с ячменем. Различный состав генов CslF , их расположение в разных видах трав и паттерны экспрессии генов могут помочь нам понять, как они эволюционировали и как они влияют на содержание (1,3; 1,4) -β-глюкана и функционируют в нем. в разное время и в разных тканях у родственных видов.Почему, например, рис содержит мало или совсем не содержит (1,3; 1,4) -β-глюкана в зерне, тогда как содержание (1,3; 1,4) -β-глюкана в зерне Brachypodium составляет до 40? % по весу, а ячмень показывает умеренное количество (1,3; 1,4) -β-глюкана с 4-10% по весу [5], [24]? Это связано с селекцией крахмалистых зерен во время одомашнивания? Ясно, что (1,3; 1,4) -β-глюкан в зерне Brachypodium в значительной степени заменяет крахмал в качестве основного запасного углевода, что согласуется с предположениями о том, что (1,3; 1,4) -β-глюкан действует как альтернативный источник метаболизируемой глюкозы в листьях молодых проростков ячменя [25].Таким образом, (1,3; 1,4) -β-глюканы у Poaceae могут играть несколько функциональных ролей в клеточных стенках и в энергетической биологии растений.
Выводы
Здесь мы охарактеризовали три недавно идентифицированных гена CslF в ячмене, которые не встречаются в рисе, Brachypodium или сорго, но присутствуют в пшенице. Хотя их участие в синтезе (1,3; 1,4) -β-глюкана еще предстоит доказать, по крайней мере два из них экспрессируются специфически в тканях листа и корня. Новые геномные данные для ячменя и родственных ему видов трав являются мощным ресурсом для характеристики эволюции и динамики сложного семейства генов CslF .В долгосрочной перспективе это будет способствовать более глубокому пониманию механизмов и процессов, регулирующих сложный углеводный обмен в стенках клеток травы.
Методы
Данные последовательности, базы данных и предварительная обработка
Доступные последовательности гена HvCslF были собраны из Национального центра биотехнологической информации (NCBI) [ HvCslF3 , GenBank: EU267179; HvCslF4 , GenBank: EU267180; HvCslF6 , GenBank: EU267181; HvCslF7 , GenBank: EU267182; HvCslF8 , GenBank: EU267183; HvCslF9 , GenBank: EU267184; и HvCslF10 , GenBank: EU267185], (http: // www.ncbi.nlm.nih.gov/). Эти последовательности использовали для поиска BLAST [26] на сборке WGS Morex ячменя, версия 3 [Международное секвенирование генома ячменя 12]. Набор данных глубокого секвенирования доступен для загрузки по адресу: http://mips.helmholtz-muenchen.de/plant/barley/index.jsp или доступен для поиска BLAST по адресу: http://webblast.ipk-gatersleben.de /ячмень/. Входные номера для CSLF последовательностей генов включены ( HvCslF3 : MLOC_59289, HvCslF4 : MLOC_74149, HvCslF6 : MLOC_57200, HvCslF7 : MLOC_51212, HvCslF8 : MLOC_52692, HvCslF9 : MLOC_59327, HvCslF10 : MLOC_13463, HvCslF11 : MLOC_19594, HvCslF12 : MLOC_7825).Кроме того, для получения генов CslF из риса был использован проект MSU по аннотации генома риса (http://rice.plantbiology.msu.edu/), а для поиска дополнительных последовательностей был проведен поиск BLAST на сборке WGS Morex ячменя. . Эти последовательности из риса и ячменя были использованы для проведения BLAST-поиска генов CslF из сорго (http://mips.helmholtz-muenchen.de/plant/sorghum/) [27], Brachypodium (http: // mips .helmholtz-muenchen.de / plant / brachypodium /) [28] и пшеница (IWGSC, URGI) [16], [17].Для поиска BLAST использовались настройки по умолчанию соответствующих веб-сайтов. Затем выравнивание проверяли на глаз, и последовательности проверяли обратным поиском BLAST в геноме риса. Идентифицированные кодирующие последовательности (за исключением пшеницы из-за доступа до публикации) приведены в наборе данных S1.
Множественное выравнивание и филогенетический / эволюционный анализ
Вышеупомянутые 79 белковых последовательностей были выровнены с использованием MUSCLE в MEGA5 [29], и на основании этого было выполнено выравнивание кодонов длиной 4323 п.н. путем замены аминокислот кодонами и одиночных аминокислотных промежутков с промежутками размером с кодон.Затем ненадежные позиции выравнивания удаляли с помощью метода BMGE [30], что приводило к выравниванию 1845 пар оснований. Последующий выбор модели и филогенетический анализ учитывали структуру кодона, имея модель нуклеотидного замещения для каждой позиции кодона: эта модель «позиции кодона», таким образом, состоит из трех моделей. Выбор модели в каждой позиции кодона был оптимизирован с использованием метода выбора модели TOPALi v2 [18], и были выбраны модели GTR + I + G для первой и третьей позиций и GTR + G для второй позиции.Затем эту модель использовали для оценки байесовского филогенетического дерева с помощью MrBayes v3.1.1 [31], запущенного из TOPALi v2. Параметры байесовского анализа составляли 2 прогона по 625 000 поколений, 25% -ное выгорание с деревьями, отобранными каждые 10 поколений, что привело к получению 100 000 деревьев из двух независимых прогонов. Значения потенциального коэффициента уменьшения масштаба (PRSF) для всех параметров были менее 1,06 (95% имели значения <1,03), что свидетельствует о хорошей сходимости (т.е. менее порогового значения PRSF, равного 1,2, как было предложено Gelman et al.[32]) двух прогонов. Дерево имело корень с десятью последовательностями из кластеров CslH и CslJ (не показаны на Фигуре 7). Апостериорные вероятности, которые показывают поддержку для каждого кластера, были преобразованы MEGA5 в проценты во время построения древовидной диаграммы, показанной на рисунке 7.
Визуальная проверка свидетельств рекомбинации была выполнена с использованием анализа по умолчанию NeighborNet в пакете SPLITSTREE [19]. Когда филогенетическая сеть NeighborNet предположила, что определенные последовательности представляют собой мозаичные последовательности, анализ был повторен, исключая их, чтобы увидеть, присутствует ли еще сигнал рекомбинации.Запуская NeighborNet в интерактивном режиме, исключая области выравнивания, было исследовано предполагаемое происхождение областей в мозаичных последовательностях. Филогенетические деревья также оценивались с использованием PhyML в TOPALi из регионов по обе стороны от предполагаемой точки останова рекомбинации, чтобы помочь в определении вероятного происхождения регионов в мозаичных последовательностях.
Пакет PAML [20] был использован для исследования вариации отношений Ka / Ks среди клад с использованием теста отношения правдоподобия для вариации давления отбора между ветвями в дереве генов на основе протокола Янга и Белявского [33].Мы обнаружили значительные различия в соотношении Ka / Ks среди клад, проверив нулевую гипотезу H 0 , согласно которой Ka / Ks было одинаковым во всех кладах (Ka / Ks равно 0,1329), по сравнению с альтернативной гипотезой H 1 . , что Ka / Ks варьировались между кладами (логарифмическая вероятность -46465,1 и -46669,5, соответственно, для H 0, и H 1, использовались для получения статистики теста отношения правдоподобия 408,4, которая была значимой при p <0,001). . Отношение Ka / Ks изменялось от 0,0398 до 0.1944 г. среди десяти клад с наибольшим количеством клад в диапазоне (0,075, 0,150).
Эксперименты по генетической локализации и последовательности РНК
Информация о генетической локализации генов и данные об экспрессии предоставлены [12]. Для генетической локализации более 3,90 гигабаз контигов последовательностей были привязаны к консенсусной генетической карте, основанной на анализе и интеграции карт ряда популяций, наибольший вклад вносила популяция рекомбинантных инбредных линий, полученных от скрещивания сортов Morex и Barke. .Восемь тканей сорта Morex подвергали РНК секвенированию с тремя повторностями на ткань [12]. Этими восемью тканями были: проросший зародыш (через четыре дня после прорастания), молодая ткань листа (от растения высотой 10 см), молодая ткань корня (от растения высотой 10 см), развивающееся соцветие (соцветие длиной 5 мм и соцветие 10-15 см). соцветие длиной мм), третье междоузлия (42-дневные растения) и две временные точки для развития зерновки (пять дней после цветения и 15 дней после цветения).Данные представлены в виде значений экспрессии FPKM (количество фрагментов на килобазу экзона на миллион картированных фрагментов).
Транзиторная
Nicotiana benthamiana экспрессионная системаПолноразмерные кДНК HvCslF11 и HvCslF12 были амплифицированы из Morex, ткани корня и молодых листьев (возрастом 10 дней) соответственно. Были использованы следующие пары праймеров: HvCslF11_F — AGCCACGGTTTACAGTACGA; HvCslF11_R — ACTACGTACGTGTCTATCCAGA; HvCslF12_F — GAAGAGCCAATGGTTTCGC; HvCslF12_R — CCAGAGAAACGGCATCATCC.Гены были клонированы в вектор входа Gateway pCR8 / GW / TOPO (Invitrogen, Карлсбад, Калифорния, США) и секвенированы на ABI 3700 (Applied Biosystems Inc., Фостер-Сити, Калифорния, США) в Австралийском исследовательском центре генома, Аделаида. , Австралия, для устранения конструкций с ошибками. В реакции рекомбинации LR вставки были перенесены в вектор-адресат шлюза pEAQ-HT-Dest1 под контролем промотора CaMW 35S [34]. В качестве положительного контроля был включен HvCslF6, а в качестве отрицательного контроля использовали вектор pEAQ-HT-Dest1 без гена устойчивости к хлорамфениколу и гена ccdB.Конструкции трансформировали в штамм AGL1 Agrobacterium tumefaciens и оставляли расти в течение ночи при 28 ° C в среде LB, содержащей рифампицин и канамицин. 2 мл ночной культуры наносили на чашку LB и выращивали в течение 2 дней при 28 ° C. 10 мл буфера для инфильтрации (10 мМ MgCl 2 , 10 мМ MES (2- (N-морфолино) этансульфоновая кислота)) добавляли на чашку, и клетки соскребали с поверхности. OD 600 измеряли и доводили до OD инфильтрации, равной 1.Добавляли 1 мкл 100 мМ ацетосирингона на мл и оставляли на 3 часа при комнатной температуре. Nicotiana benthamiana. Саженцы выращивали в теплице при 22 ° C и естественном освещении в ускорителе растений (Университет Аделаиды). Целые листья 4-недельных растений Nictotiana benthamiana инфильтрировали с нижней стороны с помощью шприца на 10 мл без иглы. Листья собирали через 6 дней, сушили вымораживанием и измельчали на шарикоподшипниковой мельнице. Анализ (1,3; 1,4) -β-D-глюкана проводили с использованием 20 мг измельченной ткани с использованием коммерчески доступных реагентов (Megazyme International Ireland Ltd, Брей, Ирландия) и протокола, основанного на [35].Модификации метода включают две промывки 50% этанолом и две промывки 100% этанолом в течение 10 минут при 97 ° C с последующей 20-минутной экстракцией при 90 ° C в 1 мл 20 мМ натрийфосфатного буфера (pH 6,5) и 1,5 часа. инкубация при 50 ° C с 40 мкл Ед / мл лихеназы. Уровни общего бета-глюкана в образцах анализировали с использованием реагента глюкозооксидаза-пероксидаза, поставляемого с набором. DP3: уровни DP4 анализировали с использованием HPAEC согласно [8] с образцами, собранными после переваривания лихеназой.
Дополнительная информация
Рисунок S1.
Проверка на микрочипе паттерна экспрессии seq РНК HvCslF11 и HvCslF12. Обработка микрочипов выполнялась на аликвотах идентичных образцов РНК, используемых для RNAseq (IBGSC, 2012 [11]), с использованием специально разработанного микрочипа Agilent для ячменя (A-MEXP-2357; www.ebi.ac.uk/arrayexpress) . Микроматрица ячменя содержит c. 61000 60-мерных зондов ячменя, полученных из предсказанных транскриптов ячменя и полноразмерных кДНК (IBGSC, 2012 [11]).Обработку проводили в соответствии с протоколом «Анализ экспрессии генов на основе одноцветной микроматрицы» (версия 6.5; Agilent Technologies). Данные были извлечены с использованием программного обеспечения Feature Extraction (FE) (v. 10.7.3.1; Agilent Technologies) с настройками по умолчанию, а затем проанализированы с использованием программного обеспечения GeneSpring GX (v. 7.3; Agilent Technologies). Данные были нормализованы с использованием одноцветных настроек Agilent FE по умолчанию в GeneSpring.
https://doi.org/10.1371/journal.pone.00
.s001
(TIF)
Рисунок S2.
Хромосомное положение членов семейства CslF подчеркивает синтению между сорго, рисом, брахиподиумом и ячменем. Рисунок был создан с использованием Strudel (см. Bayer M, Milne I, Stephen G, Shaw P, Cardle L, et al. (2011) Сравнительная визуализация генетических и физических карт с помощью Strudel. Bioinformatics 27: 1307-1308.).
https://doi.org/10.1371/journal.pone.00
.s002
(TIF)
Вклад авторов
Задумал и спроектировал эксперименты: RW CH.Проведены эксперименты: MS AL PH. Проанализированы данные: MS FW KM DM JGS GBF PH. Предоставленные реагенты / материалы / инструменты анализа: FW KM. Написал статью: MS CH RW RAB DW.
Список литературы
- 1. Управление по санитарному надзору за качеством пищевых продуктов и медикаментов (2006 г.) Маркировка пищевых продуктов: заявления о пользе для здоровья; растворимые пищевые волокна из определенных продуктов и ишемической болезни сердца. Окончательное правило. Fed Regist 71: 29248–29250.
- 2. Управление по санитарному надзору за качеством пищевых продуктов и медикаментов (2008) Маркировка пищевых продуктов: заявления о пользе для здоровья; растворимая клетчатка из определенных продуктов и риск ишемической болезни сердца.Окончательное правило. Федеральный регистр 73: 47828–47829.
- 3. Бреннан С.С., Клири Л.Дж. (2005) Потенциальное использование зерновых (1 → 3,1 → 4) -β-D-глюканов в качестве функциональных пищевых ингредиентов. J Cereal Science 42: 1–13.
- 4. Annison G (1993) Роль некрахмальных полисахаридов пшеницы в питании бройлеров. Aust J Agr Res 44: 405–422.
- 5. Бертон Р.А., Финчер Г.Б. (2012) Современные проблемы биологии клеточной стенки злаков и трав. Front Plant Sci 3: 130.
- 6.Vega-Sanchez ME, Verhertbruggen Y, Scheller HV, Ronald PC (2013) Изобилие глюкана со смешанными связями в зрелых тканях и вторичных клеточных стенках трав. Сигнальное поведение предприятия 8.
- 7. Бертон Р.А., Уилсон С.М., Хрмова М., Харви А.Дж., Ширли Н.Дж. и др. (2006) Гены, подобные синтазе целлюлозы , CslF , опосредуют синтез (1,3; 1,4) -β-D-глюканов клеточной стенки. 311: 1940–1942.
- 8. Бертон Р.А., Коллинз Х.М., Киббл Н.А.Дж., Смит Д.А., Ширли Нью-Джерси и др.(2011) Сверхэкспрессия специфических генов, подобных целлюлозосинтазе HvCslF , в трансгенном ячмене увеличивает уровни (1,3; 1,4) -β-D-глюканов в клеточной стенке и изменяет их тонкую структуру. Биотехнология растений J 9: 117–135.
- 9. Доблин М.С., Петтолино Ф.А., Уилсон С.М., Кэмпбелл Р., Бертон Р.А. и др. (2009) Ген CSLH , подобный синтазе целлюлозы ячменя , опосредует синтез (1,3; 1,4) -β-D-глюкана у трансгенного Arabidopsis . Proc Natl Acad Sci U S A 106: 5996–6001.
- 10. Доблин М.С., Петтолино Ф, Бачич А. (2010) Стенки растительных клеток: скелет растительного мира. Функциональная биология растений 37: 357–381.
- 11. Уилсон С.М., Бертон Р.А., Доблин М.С., Стоун Б.А., Ньюбигин Э.Дж. и др. (2006) Временное и пространственное появление полисахаридов стенки во время клеточности эндосперма ячменя ( Hordeum vulgare ). Planta 224: 655–667.
- 12. IBGSC (2012) Сборка физической, генетической и функциональной последовательности генома ячменя.Природа 491: 711–716.
- 13. Бертон Р.А., Джоблинг С.А., Харви А.Дж., Ширли Н.Дж., Мазер Д.Е. и др. (2008) Генетика и профили транскрипции целлюлозосинтазоподобного семейства гена HvCslF в ячмене. Физиология растений 146: 1821–1833.
- 14. Mascher M, Muehlbauer GJ, Rokhsar DS, Chapman J, Schmutz J, et al .. (2013) Закрепление и упорядочение сборок контигов NGS с помощью популяционного секвенирования (POPSEQ). Завод Дж.
- 15. Coutinho PM, Deleury E, Davies GJ, Henrissat B (2003) Развивающаяся иерархическая семейная классификация гликозилтрансфераз.J Mol Biol 328: 307–317.
- 16. Международный консорциум по секвенированию генома пшеницы. Доступно: http://www.wheatgenome.org/.
- 17. Unité de Recherche Génomique Info. Доступно: http://wheat-urgi.versailles.inra.fr/Seq-Repository] (IWGSC).
- 18. Милн I, Линднер Д., Байер М., Хусмайер Д., Макгуайр Дж. И др. (2009) TOPALi v2: богатый графический интерфейс для эволюционного анализа множественных согласований на кластерах HPC и многоядерных рабочих столах.Биоинформатика 25: 126–127.
- 19. Хусон Д.Х., Брайант Д. (2006) Применение филогенетических сетей в эволюционных исследованиях. Mol Biol Evol 23: 254–267.
- 20. Ян З. (2007) PAML 4: филогенетический анализ методом максимального правдоподобия. Mol Biol Evol 24: 1586–1591.
- 21. Han F, UIlrich SE, Chirat S, Menteur S, Jestin L, et al. (1995) Картирование локусов содержания β-глюкана и активности β-глюканазы в зерне ячменя и солода. Theor Appl Genet 91: 921–927.
- 22. Taketa S, Yuo T, Tonooka T, Tsumuraya Y, Inagaki Y, et al. (2012) Функциональная характеристика мутантов без бетаглюканов ячменя демонстрирует уникальную роль CslF6 в биосинтезе (1,3; 1,4) -β-D-глюкана. J Exp Bot 63: 381–392.
- 23. Doust A (2007) Архитектурная эволюция и ее последствия для одомашнивания трав. Энн Бот 100: 941–950.
- 24. Гийон Ф., Ларре С., Петипас Ф., Бергер А., Муссави Дж. И др. (2012) Полный обзор развития зерна у сорта Brachypodium distachyon Bd21.J Exp Bot 63: 739–755.
- 25. Roulin S, Buchala AJ, Fincher GB (2002) Индукция (1 → 3,1 → 4) -β-D-глюкангидролаз в листьях проростков ячменя, инкубированных в темноте. Планта 215: 51–59.
- 26. Альтшул С.Ф., Гиш В., Миллер В., Майерс Е. В., Липман Д. Д. (1990) Инструмент поиска базового локального выравнивания. J Mol Biol 215: 403–410.
- 27. Патерсон А.Х., Бауэрс Дж. Э., Бруггманн Р., Дубчак И., Гримвуд Дж. И др. (2009) Геном двухцветного сорго и разнообразие трав.Природа 457: 551–556.
- 28. Международная инициатива по брахиподиуму (2010) Секвенирование генома и анализ модельной травы Brachypodium distachyon. Природа 463: 763–768.
- 29. Тамура К., Петерсон Д., Петерсон Н., Стечер Г., Ней М. и др. (2011) MEGA5: Анализ молекулярной эволюционной генетики с использованием методов максимального правдоподобия, эволюционного расстояния и максимальной экономии. Mol Biol Evol 28: 2731–2739.
- 30. Criscuolo A, Gribaldo S (2010) BMGE (Block Mapping and Gathering with Entropy): новое программное обеспечение для выбора филогенетических информационных областей из нескольких выравниваний последовательностей.BMC Evol Biol 10: 210
- 31. Ronquist F, Huelsenbeck JP (2003) MrBayes 3: байесовский филогенетический вывод в смешанных моделях. Биоинформатика 19: 1572–1574.
- 32. Гельман А., Карлин Дж., Стерн Х., Рубин Д. (1995) Байесовский анализ данных. Лондон, Великобритания: Chapman & Hall.
- 33. Ян З., Белявский JP (2000) Статистические методы обнаружения молекулярной адаптации. Тенденции Ecol Evol 15: 496–503.
- 34. Sainsbury F, Thuenemann EC, Lomonossoff GP (2009) pEAQ: универсальные векторы экспрессии для легкой и быстрой временной экспрессии гетерологичных белков в растениях.Plant Biotechnol J 7: 682–693.
- 35. McCleary BV, Codd R (1991) Измерение (1-3), (1-4) -Бета-D-глюкана в ячмене и овсе — упрощенная ферментативная процедура. Журнал продовольственной науки и сельского хозяйства 55: 303–312.
5′-фланкирующая область гена гордеина B ячменя контролирует тканевую и онтогенетическую экспрессию CAT в растениях табака
Ссылки
1 апреля 1979 г. · Труды Национальной академии наук Соединенных Штатов Америки · DH Figurski , DR Helinski
15 июня 1977 г. · Журнал молекулярной биологии · PW RigbyP Berg
7 мая 1976 г. · Аналитическая биохимия · MM Bradford
1 июня 1985 г. · Cell · RD Palmiter, RL Brinster
24 апреля 1987 г. · Исследования нуклеиновых кислот · Дж. Штокхаус Л. Виллмитцер
18 января 1985 · Наука · М. Мерколак Каламе
22 августа 1985 · Природа · Дж. Ламппан Х Чуа
25 октября 1985 г. · Исследования нуклеиновых кислот · BG FordeM Kreis
ноя 26, 1984 · Исследования нуклеиновых кислот · М. Беван
1 января 1984 · Симпозиумы по количественной биологии в Колд-Спринг-Харбор · С.Л. Деллапорта Дж. Б. Хикс
1 сентября 1982 г. · Молекулярная и клеточная биология · СМ Горман Б. Ховард
1 октября, 1982 · Ячейка · H Гиллей К. Э. Ричардс
1 мая 1985 г. · Труды Национальной академии наук Соединенных Штатов Америки · К. Сенгупта-Гопалан Дж. Д. Кемп
1 сентября 1985 г. · Физиология растений · Дж. С. Гринвуд, М. Дж. Криспилс
30 мая, 1986 · Наука · R FluhrN H Chua
5 июня 1987 · Наука · R KayJ McPherson
Цитаты
1 января 1990 · Молекулярная биология растений · DJ LeisyT W Okita
1 ноября 1990 · Молекулярная биология растений Биология · Г. А. Томпсон Б. А. Ларкинс
1 января 1991 г. · Молекулярная биология растений · Ф Такайва Като
1 ноября 1993 г. · Молекулярная биология растений М. Х. Белл Д. Фрэнсис
1 июля 1989 г. · Молекулярная биология растений · Б. Ли М. Джонс
1 декабря 1991 г. · Молекулярная биология растений · Дж. Энтвистл В. Камерон-Миллс
1 марта 1990 г. · Молекулярная биология растений · А.Дж. Мацке М. А. Мацке
1 сентября 1993 г. · Молекулярная биология растений · Р. Мантейфель, Р. Паниц
1 ноября , 1991 · Молекулярная биология растений · GA HullP R Shewry
90 002 1 октября 1994 г. · Молекулярная биология растений · R SchubertH Bäumlein1 марта 1989 г. · Молекулярная биология растений · RT DeroseT C Hall
1 июля 1990 г. · Молекулярная и общая генетика: MGG · A PeterhansJ Paszkowski
1 мая, 1993 · Трансгенные исследования · X Zhou, YL Fan
, 1 января 1988 г. · Carlsberg Research Communications · J Entwistle
23 мая 2007 г. · Отчеты о растительных клетках · Xiaoping ChenGuoying Wang
27 октября 2009 г. · Genetica
1 апреля 2010 г. · Молекулярная биология растений · Ариадна ПеремартиТереза Капелл
10 ноября 1998 г. · Журнал растений: клеточная и молекулярная биология · M MenaP Carbonero
28 мая 2008 г. · Журнал биотехнологии растений · Agnelo FurtadoFumio Takaiwa
января 27, 1989 · Cell · RB GoldbergL Perez-Grau
1 апреля 1989 · BioEssays: Новости и обзоры в молекулярной, клеточной биологии и биологии развития · GA Thompson, BA Larkins
5 февраля 2015 · Физиология и молекулярная биология растений: Международный Журнал функциональной биологии растений · J Beslin JoshiD Sudhakar
10 ноября 1998 г. · The Plant Journal: для клеточной и молекулярной биологии · P MarzábalM Torrent
26 марта 2002 г. · Journal of Experimental Botany · Питер Р. Шури, Найджел Г. Хэлфорд
27 октября 2017 г. · Журнал экспериментальной ботаники · Хайко Вайхерт · Ханс Вебер
1 августа 1996 г. · Молекулярная биология растений · J RoyoP Carbonero
20 сентября 1995 г. · Молекулярная и общая генетика: MGG · I DiazP Carbonero
15 августа, 2002 · Journal of Biosciences · J Baird Callicott
15 августа 1994 г. · Молекулярная и общая генетика: MGG · P GallusciR D Thompson
1 июня 1990 г. · Молекулярная и общая генетика: MGG · MA BogueT L Thomas
1 августа, 1991 · Молекулярная и общая генетика: MGG · SS KlemsdalO A Olsen
1 мая 1990 · Молекулярная и общая генетика: MGG · P GuercheG Pelletier
Произошла ошибка при настройке вашего пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности.Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки вашего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались.Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie.Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
точное картирование локуса восприимчивости к чистым пятнам хромосомы ячменя 6H | G3: Гены | Геномы
Реферат
Сетчатая пятнистость, вызванная некротрофным грибковым патогеном Pyrenophora teres f. teres — это деструктивное заболевание листьев ячменя, способное вызвать значительную потерю урожая в основных производственных регионах по всему миру. Сложность генетических взаимодействий паразит-хозяин в этой патосистеме препятствует развертыванию эффективной устойчивости у сортов ячменя, что требует более глубокого понимания взаимодействий. Здесь мы сообщаем о картировании с высоким разрешением доминантного локуса восприимчивости вблизи центромеры хромосомы 6H у сортов ячменя Rika и Kombar, которые предположительно являются мишенью некротрофных эффекторов из P.teres f. teres изолирует 6A и 15A соответственно. Использование изолятов потомства, полученных от скрещивания P. teres f. teres изолятов 6A × 15A, несущих одиночные главные локусы вирулентности ( VK1 , VK2 и VR2 ), позволили провести менделизацию единичных обратных взаимодействий ген-ген в популяции с высоким разрешением, состоящей из 2976 Rika × Комбар-рекомбинантные гаметы. Brachypodium distachyon synteny использовали для развития и насыщения области чувствительности маркерами, ограничивая ее до ∼0.24 см и была построена частичная физическая карта. Эта генетическая и физическая характеристика дополнительно разрешила доминантный локус восприимчивости, обозначенный Spt1 (восприимчивость к P. teres f. teres ). Картирование с высоким разрешением и косегрегация гена / ов Spt1.R и Spt1.K указывает на тесно связанные гены в отталкивании или аллели, возможно, нацеленные на различные некротрофные эффекторы. Недавно разработанные геномные ресурсы ячменя значительно повышают эффективность усилий по позиционному клонированию ячменя, что продемонстрировано точным картированием Spt1 и физической идентификацией контигов, о которых сообщается здесь.
Некротрофный гриб Pyrenophora teres является возбудителем сетчатой пятнистости ячменя ( Hordeum vulgare ) и существует в двух формах, которые, хотя морфологически неразличимы под микроскопом, могут быть дифференцированы по симптомам, наблюдаемым на чувствительных генотипах ячменя ( Маклин, и др., , 2009; Смедегард-Петерсон, 1971). Сетчатая пятнистость (NFNB), вызываемая P. teres f. teres проявляется некротическими поражениями с поперечными и продольными бороздками, образующими сетчатую структуру некроза, часто сопровождающуюся хлорозом (Mathre 1997).Пятно болезни представляет собой сетчатую пятнистость (SFNB), вызываемую P. teres f. maculata вызывает эллиптические некротические поражения, окруженные хлорозом (McLean et al. 2009; Smedegård-Peterson 1971). И NFNB, и SFNB являются деструктивными заболеваниями листьев, которые могут вызывать потери урожая на 10-40% (Mathre 1997; Murray and Brennan 2010) и снижение качества конечного использования ячменя. Хотя аналогичные применения фунгицидов и культурные практики могут использоваться для борьбы с этими заболеваниями, предпочтительным методом лечения является резистентность хозяина (Mathre 1997).Однако, несмотря на то, что эти два патогена тесно связаны и генетически очень похожи, их генетические взаимодействия между хозяином и патогеном различны (обзор Liu et al. 2011), поэтому при развертывании генетической устойчивости к ним следует относиться как к различным заболеваниям.
Использование глобально разнообразных сортов ячменя и P. teres f. teres изолятов, несколько генетических исследований позиционировали как доминантную, так и рецессивную устойчивость к NFNB в центромерной области хромосомы 6H ячменя, предполагая наличие множества разнообразных генов устойчивости или восприимчивости.Предполагается, что эти гены-хозяева взаимодействуют с разными P. teres f. teres эффекторов / белков авирулентности из различных изолятов, приводящих к дифференциально совместимым или несовместимым реакциям, наблюдаемым в этой патосистеме (Ho et al. 1996; Steffenson et al. 1996; Raman et al. 2003; Cakir et al. al. 2003; Ma et al. 2004; Manninen et al. 2006; Friesen et al. 2006a; Abu Qamar et al. 2008; St. Pierre et al. 2010). Абу Камар и др. (2008) картировал два рецессивных гена устойчивости, которые существуют при отталкивании, в популяции с двойным гаплоидом, полученной от скрещивания линий ячменя Rika и Kombar. Рика устойчива к P. teres f. teres изолят 15А и восприимчивы к P. teres f. teres изолят 6A, тогда как Kombar устойчив к P. teres f. teres изолят 6А и восприимчив к P.teres f. teres изолятор 15А. Локусы восприимчивости были картированы в области ~ 5,9 сМ вблизи центромеры на хромосоме 6H ячменя. Далее область была ограничена интервалом ~ 3,3 сМ с помощью маркеров на основе EST (Liu et al. 2010).
Комплексный генетический анализ с использованием популяции двух родителей P. teres f. teres. изолятов 6A и 15A, которые проявляют реципрокную вирулентность в отношении Rika и Kombar, соответственно, идентифицировали четыре вирулентных QTL в патогене.Два QTL, обозначенные как VR1 и VR2 для вирулентности на Rika 1 и 2, были идентифицированы для P. teres f. teres , изолят 6A и два QTL, VK1 и VK2 , для вирулентности на Kombar 1 и 2, были идентифицированы для P. teres f. teres изолят 15А (Shjerve et al. 2014). Было высказано предположение, что в основе этих вирулентных QTL лежат селективные токсины хозяина или некротрофные эффекторы, поскольку недавно в этой патосистеме было показано, что белковый эффектор из P.teres f. teres Изолят 0-1, обозначенный PttNE1, взаимодействует с единственным геном доминантной чувствительности на хромосоме 6H у ячменя cv. Hector, индуцирующий некротрофную эффекторную триггерную восприимчивость (NETS) (Liu et al. 2015). Кроме того, другие близкородственные грибы-некротрофные аскомицеты, такие как Pyrenophora tritici-repentis и Parastagonospora nodorum , продуцируют некротрофные эффекторы, которые изменяют ответы врожденного иммунитета хозяина, что приводит к усилению заболевания (Tomas et al. 1990; Стрелков и др. 1999; Лю и др. 2009, 2012; Gao et al. 2015). Белковый некротрофный эффектор ToxA, продуцируемый P. tritici-repentis (Ptr ToxA) и P. nodorum (SnToxA), взаимодействует с доминирующим белком чувствительности хозяина Tsn1 (Friesen et al. 2006b; Faris et al. 2010). Tsn1 имеет белковые домены, подобные типичным белкам устойчивости, которые придают устойчивость к биотрофным патогенам за счет индукции строго регулируемых путей запрограммированной гибели клеток (PCD).Обычно считается, что эти пути эволюционировали для изоляции биотрофных патогенов, которым требуются живые ткани хозяина для завершения своего жизненного цикла. Несоответствующее проявление этих врожденных путей резистентности посредством использования некротрофных эффекторов некротрофными патогенами позволяет им разрушать и использовать пути иммунитета хозяина для инфицирования и завершения своего жизненного цикла в хозяине. Таким образом, они используют некротрофные эффекторы для нацеливания на специфические гены восприимчивости хозяина, которые иначе представляют собой иммунные рецепторы, чтобы вызвать NETS.Эти взаимодействия между хозяином и патогеном аналогичны классической парадигме ген-ген (Flor 1956), предложенной Г. Н. Флор (Flor 1956), но считаются обратными отношениями ген-ген, в которых специфические взаимодействия, приводящие к ПКС, приводят к совместимому взаимодействию. (восприимчивость) количественно в зависимости от характера образа жизни патогена (Friesen et al. 2007). Однако также представляется, что в этой патосистеме также существуют классические генетические или доминантные резистентности, как Friesen et al. (2006a) идентифицировал доминирующую устойчивость к трем различным изолятам NFNB, которые, что интересно, также картировались в центромерной области хромосомы 6H ячменя.
Виды трав, такие как глобально важные зерновые культуры ячмень, пшеница ( Triticum aestivum ), кукуруза ( Zea mays ), рис ( Oryza sativa ) и сорго ( Sorghum bicolor ), отличались от обычного предок ∼55–70 миллионов лет назад (Kellogg 2001). На протяжении всей их эволюционной истории было показано, что относительное содержание генов и порядок сохраняются между видами трав, при этом большая часть изменения размера генома происходит из-за увеличения повторяющихся элементов (Bennetzen and Freeling 1997; Kellogg 2001).Хотя наблюдаются разрывы синтении, приписываемые вставкам и транслокациям хромосом, общий порядок генов хорошо сохраняется (IBI 2010). Секвенирование и аннотация генома Brachypodium distachyon предоставили полезный инструмент для открытия генов у других видов трав, особенно видов сельскохозяйственных культур с большими геномами в результате распространения повторяющихся элементов ДНК. Эти большие повторяющиеся геномы усложняют сборку геномных последовательностей, так что имеется адекватная сборка пространств генов с низкой копией, но много пробелов в повторяющихся областях, поэтому надежные сборки генома не были достигнуты (Mur et al. 2011; IBGSC 2012). Сохранение синтения между последовательностью модельного генома травы и ячменя и пшеницы ускоряет разработку маркеров в геномах крупных трав, обеспечивая быстрое насыщение регионов для позиционного клонирования (IBI 2010; Mur et al. 2011)
Проект сборки пространство генов ячменя было недавно завершено, однако почти полная сборка генома размером ~ 5,1 ГБ, который состоит примерно из 84% повторяющихся элементов, является сложной задачей с текущим биоинформатическим программным обеспечением, но предварительная физическая карта генома ячменя был собран с помощью высокоинформативного фингерпринта клонов бактериальной искусственной хромосомы (ВАС) и включения последовательности полногеномного дробовика (WGS), концевой последовательности ВАС (BES), последовательности из выбранных клонов ВАС и интеграции данных генетического картирования (IBGSC). 2012).Кроме того, дополнительное закрепление контигов WGS было выполнено посредством секвенирования популяции (POPSEQ), и клоны BAC были собраны в минимальные мозаичные пути через собранные контиги (Mascher et al. 2013; Ariyadasa et al. 2014). Кроме того, 15 622 cv. BAC Morex были секвенированы и впоследствии интегрированы с консенсусной картой и анализом синтении в легкодоступное программное обеспечение HarvEST (Muñoz-Amatriaín et al. 2015). Использование этого ресурса позволяет выполнять физическое картирование регионов, идентифицированных стратегиями клонирования на основе генетических карт, что способствует более эффективной идентификации генов-кандидатов.
Доминантные гены предрасположенности Spt1.R / Spt1.K (ранее rpt.k / rpt.r ), которые, как предполагается, придают доминантную восприимчивость к P. teres f. teres изоляты 6А и 15А, по-видимому, действуют сложным образом. Мы предполагаем, что аллели Spt1 , Spt1.R и Spt1.K или нескольких тесно связанных генов придают основной эффект доминантной чувствительности к P. teres f. teres изолирует 6A и 15A соответственно.Здесь мы описываем картирование с высоким разрешением доминантного локуса восприимчивости, расположенного в центромерной области хромосомы 6H ячменя. Картирование было выполнено с использованием маркеров, разработанных с использованием синтенических отношений с хромосомой B. distachyon 3. Кроме того, идентификация физических контигов ячменя, обеспеченная недавно разработанными геномными ресурсами ячменя, была использована для построения частичной физической карты область. Результаты этого исследования вскоре облегчат идентификацию гена доминантной восприимчивости Spt1 и позволят провести последующий функциональный анализ его аллелей, функционирование которых предположительно с помощью NETS.Это исследование заполнит пробелы в знаниях, касающихся этих сложных генетических взаимодействий паразит-хозяин и лежащих в основе молекулярных механизмов, определяющих совместимые взаимодействия в этой сложной патосистеме ячменя и NFNB.
Материалы и методы
Биологические материалы
Два P. teres f. teres изолятов 6A и 15A, а также потомство, идентифицированное как содержащее одиночный QTL вирулентности для VK1, VK2 и VR2 (Shjerve et al. 2014) от скрещивания изолятов 6A и 15A, были использованы для фенотипирования.Модель P. teres f. teres Изолят 6А был собран в округе Фресно, Калифорния (Wu et al. , 2003), а изолят 15А был собран в округе Солано, Калифорния (Стеффенсон и Вебстер, 1992).
Скандинавский сорт двурядного ячменя Rika и сорт шестирядного ячменя США Комбар ранее были идентифицированы как имеющие взаимную реакцию на P. teres f. teres изолирует 6A и 15A. Рика восприимчива к P. teres f. teres изолят 6A и устойчив к P. teres f. teres изолятор 15А. Комбар проявляет противоположную реакцию, будучи восприимчивым к P. teres f. teres изолят 15A и устойчив к P. teres f. teres изолят 6А (Abu Qamar et al. 2008). Rika и Kombar были скрещены и продвинуты для создания популяции из 1488 F 2 особей, представляющих 2976 гамет, которые впоследствии были генотипированы для идентификации рекомбинантных гамет в области Spt1 .Ранее идентифицированное критическое рекомбинантное потомство двойных гаплоидов (DH), идентифицированное из популяции, состоящей из 118 особей, полученных от скрещивания Rika и Kombar (Abu Qamar et al. 2008), также было генотипировано для проверки рекомбинантов DH в области NFNB Spt1 . , который ранее назывался рецессивными генами устойчивости rpt.k и rpt.r .
Генотипирование и идентификация рекомбинантных гамет
Два ранее идентифицированных маркера фланкирующих простых повторов последовательности (SSR), Bmag0173 на дистальной стороне и Rbah31g15 на проксимальной стороне (Liu et al. 2010), были использованы для генотипирования 1488 Rika × Kombar F 2 особей. Выделение ДНК для первоначального генотипирования F 2 было выполнено в центре генотипирования зерновых Министерства сельского хозяйства и сельскохозяйственных исследований США (USDA-ARS), Фарго, Северная Дакота, с использованием высокопроизводительной процедуры, описанной Tsilo et al. (2009).
После того, как F 2 особей с рекомбинацией в области Spt1 были идентифицированы на начальном этапе генотипирования с помощью маркеров SSR на секвенаторе ABI, рекомбинантные проростки пересаживали из контейнеров в 6-дюймовые горшки, выращенные в теплице. и позволили самостоятельно произвести семена F 2: 3 .Пятнадцать растений F 2: 3 из каждой семьи выращивали в теплице, и геномную ДНК выделяли из образцов ткани листьев длиной приблизительно 2 см, собранных в пробирки Эппендорфа на 1,5 мл. Ткань листа гомогенизировали в 400 мкл буфера для экстракции (200 мМ Трис-HCl pH 7,5, 250 мМ NaCl, 25 мМ EDTA, 0,5% SDS) с помощью одноразового измельчителя тканей с последующим добавлением 200 мкл хлороформа. Образцы встряхивали в течение 10 секунд и центрифугировали при 15 600 × g в течение 10 минут. Супернатант (~ 300 мкл) переносили в новую 1.Добавляли 5 мл пробирку и 300 мкл изопропанола. Образцы перемешивали путем переворачивания и инкубировали при комнатной температуре в течение 5 минут с последующим центрифугированием при 15,600 × g в течение 10 минут. Гранулы промывали 70% этанолом, сушили и ресуспендировали в 100 мкл ddH 2 О. 15 особей F 2: 3 из каждого семейства F 2 , идентифицированных с критической рекомбинацией в области Spt1 . были генотипированы с использованием кодоминантного маркера rpt-M4 для идентификации гомозиготных рекомбинантных индивидуумов, которые представляют собой бессмертные критические рекомбинанты (ICR).Особи ICR получили обозначение потомства и использовались для дальнейшего фенотипирования и генотипирования.
Разработка маркеров STS / SNP
Используя синтенную взаимосвязь между хромосомой 6H ячменя и хромосомой 3 B. distachyon , были разработаны генетические маркеры для насыщения области Spt1 с использованием аннотированной последовательности генома B. distachyon . (IBI 2010). Ортологические локусы в B. distachyon , соответствующие дистальному фланкирующему маркеру EST BE636841 и проксимальному маркеру SSR Rbah31g15, были использованы для идентификации ∼1.02 Mb синтенической последовательности геномной ДНК B. distachyon . Метки экспрессируемой последовательности (EST) ортологичного ячменя были идентифицированы с использованием поиска Blastn, ограниченного EST other_ Hordeum vulgare , которые соответствуют генам B. distachyon в данном регионе. Набор этих EST был выбран с интервалами ~ 60 т.п.н. с использованием прогнозируемой функции в качестве критерия отбора, поскольку некоторые гены имеют более высокую вероятность содержания полиморфизмов по сравнению с другими. Например, гены NBS-LRR обычно имеют большее разнообразие, чем высококонсервативные гены домашнего хозяйства.Олигонуклеотидные праймеры были сконструированы из последовательностей EST ячменя и использованы для получения ампликонов из геномной ДНК Rika и Kombar с помощью ПЦР (таблица 1). Геномную ДНК экстрагировали, как описано ранее. Параметры реакций ПЦР были следующими: 94 ° в течение 5 минут, затем 35 циклов по 94 ° в течение 30 секунд, 62 ° в течение 30 секунд и 72 ° в течение 60 секунд, с последующим окончательным удлинением при 72 ° в течение 7 секунд. мин. Реакции ПЦР включали 1,25 единиц стандартной полимеразы Taq NEB, прямого и обратного праймеров (1,2 мкМ), стандартного буфера Taq NEB (1 ×) и dNTP (200 мкМ) в реакциях по 25 мкл.Продукты ПЦР визуализировали на 1% агарозном геле, содержащем GelRed (Biotium, Калифорния), а затем очищали с использованием E.Z.N.A. Cycle Pure Kit (Omega Bio-Tek, Norcross, GA) в соответствии со стандартным протоколом производителя. Ампликоны были секвенированы (McLab и Genscript), и однонуклеотидные полиморфизмы (SNP) были идентифицированы путем выравнивания с использованием программного обеспечения Vector NTI (Invitrogen) и инструментов выравнивания нуклеотидных последовательностей NCBI. Маркеры сайтов, меченных последовательностью (STS), были разработаны на основе SNP путем конструирования специфичных для генотипа праймеров со специфичными для SNP нуклеотидами, соответствующими 3′-концевому нуклеотиду праймеров (таблица 1).Если устойчивый маркер STS не удалось разработать, критические рекомбинанты амплифицировали с использованием исходных пар праймеров и ампликоны секвенировали по Сэнгеру (Genscript).
Таблица 1 Последовательности праймеров (5’– 3 ‘) для полиморфных маркеров, полученных из ортологов ячменя генов Brachypodium distachyon в области Spt1Подготовка библиотеки PCR-GBS и ионное торрент-секвенирование
Генотипирование с помощью ПЦР путем секвенирования Также была создана панель (PCR-GBS), состоящая из двух ранее разработанных маркеров SNP из базы данных T3 (https: // triticeaetoolbox.org / barley /) и 11 пар праймеров, разработанных на основе предсказанной кодирующей, интронной или 3′-UTR-последовательности из ортологов ячменя генов B. distachyon в пределах области с целью обнаружения SNP. Кроме того, четыре ранее разработанных маркера STS были включены в панель для регенотипирования и внутреннего контроля. Адаптер CS1 из 22 нуклеотидов (5′-ACACTGACGACATGGTTCTACA-3 ‘) (Fluidigm) был добавлен к 5′-концу прямых праймеров, а адаптер CS2 из 22 нуклеотидов (5′-TACGGTAGCAGAGACTTGGTCT-3′) (Fluidigm) был добавлен к 5’-концу. ‘Конец обратных праймеров.Были сконструированы адаптерные праймеры со штрих-кодом, содержащие адаптерную последовательность Ion Torrent A, уникальный 12-нуклеотидный штрих-код и адаптерную последовательность CS1 (5’-CCATCTCATCCCTGCGTGTCTCCGACTCAG (NNNNNNNNCGAT) ACACTGACGACATGGTTCTACA-3 ‘). Был разработан универсальный обратный праймер, содержащий последовательность адаптера Ion Torrent P1 и последовательность адаптера CS2 (5’-CCACTACGCCTCCGCTTTCCTCTCTATGGGCAGTCGGTGATTACGGTAGCAGAGACTTGGTCT-3 ‘). Праймеры мультиплексировали, добавляя по 5 мкл каждого прямого и обратного праймера (100 мкМ) в стерильный 1.Пробирка Эппендорфа 5 мл и 820 мкл свободной от нуклеаз H 2 O для конечного объема пула праймеров 1 мл. Реакции ПЦР включали 1,5 мкл геномной ДНК, 1 мкл пула праймеров (100 нМ каждый праймер) и 2,5 мкл Platinum Multiplex PCR Master Mix (Life Technologies). Концентрации ДНК были нормализованы до ~ 20 нг / мкл, и 1,5 мкл каждого образца было внесено в отдельные лунки 96-луночного планшета. Копии Rika, Kombar, RK107, RK121 и RK122 также были включены в пластину. Первичную реакцию ПЦР проводили следующим образом: начальная денатурация при 94 ° в течение 10 мин, 10 циклов при 94 ° в течение 20 секунд и 64 ° с уменьшением каждого цикла на 0.8 ° в течение 60 секунд, затем 20 циклов по 94 ° в течение 20 секунд, 57 ° в течение 60 секунд и 68 ° в течение 30 секунд, заканчивая окончательным расширением 72 ° в течение 3 минут. Затем планшет для ПЦР ненадолго центрифугировали и к каждому образцу добавляли 15 мкл свободной от нуклеаз H 2 O. Затем планшет герметично закрывали, встряхивали, недолго центрифугировали и аликвотировали по 2 мкл каждого образца в новый 96-луночный планшет. Кроме того, 2 мкл каждой реакции разделяли на 1% агарозном геле, содержащем GelRed, для обеспечения успешных реакций. Реакция ПЦР со штрих-кодированием состояла из 0.235 мкл H 2 O, 0,625 мкл стандартного буфера Taq NEB (1,25 ×), 0,1 мкл dNTP (500 мкМ), 1 мкл универсального обратного праймера (1 мкМ), 1 мкл праймера адаптера уникального штрих-кода (0,4 мкМ), 0,04 мкл полимеразы NEB Taq (0,2 единицы) и 2 мкл матрицы (разбавленная первичная реакция ПЦР). Параметры ПЦР такие же, как у первичной реакции. После ПЦР планшет ненадолго центрифугировали и каждый образец разбавляли, добавляя 15 мкл H 2 O. Образцы объединяли путем аликвотирования 5 мкл каждого образца в 1.Пробирка Эппендорфа 5 мл и очищена с использованием E.Z.N.A. Набор Cycle Pure. Затем объединенную библиотеку амплифицировали для подготовки к секвенированию. Реакции ПЦР включали 2 мкл библиотеки секвенирования, 13 мкл H 2 O, 2,5 мкл стандартного буфера Taq NEB (1 ×), 3 мкл праймера ABC1 (0,6 мкМ), 3 мкл праймера P1 (0,6 мкМ), 0,5 мкл дНТФ. (200 мкМ) и 1 мкл полимеразы NEB Taq (5 единиц). Параметры ПЦР были следующими: начальная денатурация при 95 ° в течение 5 минут, затем 8 циклов 94 ° в течение 30 секунд, 62 ° в течение 30 секунд и 72 ° в течение 30 секунд, завершение окончательным удлинением при 72 ° в течение 7 минут. .Амплифицированную библиотеку количественно оценивали с использованием набора Qubit dsDNA High Sensitivity Kit и Qubit Fluorometer (Life Technologies) и разбавляли до ~ 3 пг / мкл. Секвенирование проводили на Ion Torrent PGM с использованием набора Ion PGM Template OT2 200, набора Ion PGM Sequencing 200 v2 и чипа Ion 314 (Life Technologies).
Вызов SNP
Считывания секвенирования были обрезаны по 22 нуклеотидам на 5′- и 3′-концах для удаления адаптеров ПЦР. Обрезанные чтения были сопоставлены с эталонным файлом, состоящим из ранее идентифицированных последовательностей ампликонов из базы данных T3, ортологичной последовательности EST ячменя, соответствующей выбранному B.distachyon в этом регионе и последовательности из ранее разработанных маркеров STS с использованием алгоритма BWA-MEM с настройками по умолчанию (Li and Durbin 2009), и выравнивания были преобразованы в файлы BAM с помощью SAMtools (Li et al. 2009). SNP были идентифицированы с помощью инструмента Unified Genotyper инструментария Genome Analysis Toolkit (DePristo et al. 2011). Затем названные SNP были отфильтрованы на основе параметров минимального качества генотипа 10 и минимальной глубины чтения на сайт для каждого человека 3.
Фенотипирование
ICR были фенотипированы с помощью P. teres f. teres изолятов 6A, 15A и потомства изолятов 15 × 6A # 20, 15A × 6A # 63 и 15A × 6A # 72 с менделизированной вирулентностью QTL VK1 , VK2 и VR2 , соответственно . Фенотипирование проводили, как описано Friesen et al. (2006a). Вкратце, инокулят получали путем посева пробок из агарозы из P. teres f. teres изолятов на среде V8 PDA (150 мл сока V8, 10 г картофельного агара с декстрозой, 10 г агарозы, 3 г CaCO 3 и 850 мл H 2 O).Культуры грибов инкубировали при комнатной температуре в темноте в течение 5 дней, помещали на свет при комнатной температуре на 24 часа, а затем возвращали в темноту при 15 ° С на 24 часа. Конидии собирали путем заливки планшетов H 2 O и соскабливания культуры с помощью стерильной петли для посева. Инокулят доводили до концентрации ~ 2000 спор / мл и добавляли одну каплю Твина 20 на 50 мл инокулята. По три семени каждой линии ICR высаживали в один контейнер (Stuewe and Sons, Inc., Corvallis, OR.) И три контейнера были посажены для каждого ICR, всего девять сеянцев на линию. Сеянцы выращивали в тепличных условиях до полного раскрытия второго листа, примерно через 14 дней. Инокулят наносили, как описано Friesen et al. (2006a) и инкубировали при 21 ° и 100% относительной влажности в течение 22–24 часов. Затем растения перемещали в камеру для выращивания при 21 ° с 12-часовым фотопериодом и оценивали через 7 дней после инокуляции по шкале от 1 до 10 (Tekauz 1985). Девять растений на линию оценивали вместе как одну экспериментальную единицу, и посевы повторяли трижды.
Построение карты с высоким разрешением
Всего 14 ICR, содержащих рекомбинацию между фланкирующими маркерами Bmag0173 и Rbah31g15, были идентифицированы и использованы при построении карты с высоким разрешением. Генетические расстояния были рассчитаны на основе частоты рекомбинации между маркерами и обозначены как единицы карты (M.U.).
Ячмень iSELECT 9K chip
Ячмень 9k Illumina Infinium iSELECT Анализ был использован для генотипирования родителей Rika и Kombar и восьми линий ICR (RK122, 125, 130, 139, 151, 173, 183 и 188) с рекомбинацией в области Spt1 .Генотипирование проводили, как описано Muñoz-Amatriaín et al. (2014).
Скрининг библиотеки ВАС и построение физической карты
Использование текущей физической карты ячменя (IBGSC 2012), содержащей закрепленные последовательности WGS и ВАС, а также библиотеки ВАС (MTP) Morex 6H (Mascher et al. 2013) ; Ariyadasa et al. 2014), генетические маркеры с карты высокого разрешения были привязаны к физическим контигам. Последовательности маркеров внутри региона использовались при поиске BLAST в базе данных Morex WGS для идентификации положительных контигов WGS (http: // webblast.ipk-gatersleben.de/barley/viroblast.php). Когда возможно, были идентифицированы положения контигов WGS на более крупных контигах с отпечатками пальцев (FPC). Те маркеры, которые не давали заякоренных контигов WGS, использовали для скрининга библиотеки Morex 6H MTP BAC с помощью ПЦР (INRA-CNRGV) для определения ее положения на FPC. Кроме того, библиотека Morex ВАС (Yu et al. 2000) была подвергнута скринингу с использованием маркеров rpt-M8, rpt-M12 и rpt-M13, как описано Brueggeman et al. (2002). Положительно идентифицированные клоны ВАС высевали на планшеты LB, содержащие хлорамфеникол (50 мкг / мл), и инкубировали в течение ночи при 37 °.Затем колонии инокулировали в жидкий LB, содержащий хлорамфеникол (50 мкг / мл), и инкубировали в течение 16 часов при 37 ° при встряхивании при 230 об / мин. ДНК ВАС выделяли, как описано в Brueggeman et al. (2002). ПЦР проводили на ДНК ВАС с маркерами rpt-M8, rpt-M12 и rpt-M13 для идентификации соответствующих клонов ВАС с использованием вышеупомянутого протокола ПЦР. Кроме того, база данных HarvEST, в которой хранятся последовательности ВАС из публикации Yu et al. (2000), был проведен поиск BLAST с использованием маркерных последовательностей для идентификации и подтверждения дополнительных клонов ВАС (Muñoz-Amatriaín et al. 2015).
Секвенирование и сборка библиотеки ВАС ДНКиз выбранных ВАС 783N21 и 650G13 (Yu et al. 2000) выделяли, как описано ранее. ДНК ВАС была объединена и секвенирована в Центре молекулярной биологии и геномики Университета штата Вашингтон с использованием технологии Ion Torrent PGM. Считывания секвенирования были de novo , собранными с использованием CLC Bio Genomics Workbench (Qiagen), требующего 95% гомологии последовательностей и минимального размера контига 1 т.п.н. для сборки.
Доступность данных
Данные и реактивы доступны по запросу. Дополнительный материал, файл S1 содержит генотипы ICR из недавно разработанных маркеров. Файл S2 содержит необработанные фенотипические данные для Rika, Kombar и ICR от прививок вышеупомянутым P. teres f. teres изолятов.
Результаты
Первоначальное генотипирование и фенотипирование критических рекомбинантов
Всего 1488 F 2 особей и 118 линий DH, полученных от скрещивания Rika × Kombar, были генотипированы фланкирующими SSR-маркерами Bmag0173 и Rbah31g15.Было идентифицировано тринадцать F 2 индивидуумов и одна линия DH, которые содержали рекомбинацию в пределах ранее ограниченной области rpt.r / rpt.k , теперь называемой областью Spt1 . Впоследствии семейства F 2: 3 этих 13 критических рекомбинантов были генотипированы кодоминантным маркером rpt-M4, и для каждого семейства F 2: 3 был идентифицирован бессмертный критический рекомбинантный (ICR) индивидуум.
Фенотипирование линий ICR
Индивидуумов из каждого семейства ICR и одинокий особь DH затем фенотипировали с помощью Pyrenophora teres f. teres изолирует 15A, 6A, 15 × 6A # 20, 15A × 6A # 63 и 15A × 6A # 72. Линии, в которых средняя реакция на заболевание составляла пять или меньше, были классифицированы как устойчивые, а линии, в которых средняя реакция на заболевание превышала пять, считались восприимчивыми. После инокуляции изолятом 15А, Рика и Комбар показали средние реакции 1,67 и 7,5 соответственно. В общей сложности пять ICR были классифицированы как устойчивые с индивидуальным средним значением от 1 до 3,33 и общим средним значением 1.77. Остальные девять ICR показали индивидуальные средние реакции на заболевание в диапазоне от 6,83 до 7,83 с общим средним значением 7,59 и были классифицированы как восприимчивые (таблица 2). При инокулировании изолятом потомства 15A × 6A # 20 (Shjerve et al. 2014), содержащим только локус вирулентности VK1 , Рика и Комбар показали средние реакции заболевания 1,83 и 6,83 соответственно. Пять ICR были признаны устойчивыми, а девять — восприимчивыми. Средняя реакция на заболевание устойчивых линий колебалась от 1.83–3,83 при среднем среднем 2,73. Для восприимчивой группы индивидуальные средние значения варьировались от 6,67 до 8,83 с общим средним значением 7,74 (таблица 2). Фенотипические данные инокуляции вторым изолятом потомства, 15A × 6A # 63, несущим только локус вирулентности VK2 (Shjerve et al. 2014), показали ту же картину. Средняя реакция на заболевание для Рики и Комбара составила 2,17 и 7,17 соответственно. Ранее классифицированные устойчивые и чувствительные ICR снова сгруппированы вместе.Средние баллы по шкале болезней устойчивых людей варьировались от 1,5 до 2 с общим средним значением 1,77. У восприимчивых людей средний балл по заболеванию варьировал от 5,33 до 7,17, а общее среднее значение — 6,35 (таблица 2).
Таблица 2 Средние показатели заболевания для Rika, Kombar и устойчивых / чувствительных ICR с изолятами PttПосле инокуляции ICR с изолятом 6A, Rika и Kombar показали типы реакций 9,5 и 4,17, соответственно. Среднее количество типов реакции на заболевание для шести устойчивых линий колебалось от 3.5–5 с общим средним значением 4,06. В общей сложности восемь человек были признаны восприимчивыми, и средний тип реакции варьировался от 5,50 до 9,33 с общим средним значением 7,65 (таблица 2). После инокуляции изолятом потомства 15A × 6A # 72, несущим только локус вирулентности VR2 (Shjerve et al. 2014), Рика и Комбар показали средние типы реакции 7,83 и 1, соответственно. В общей сложности девять ICR были признаны устойчивыми со средним баллом по заболеванию от 1,00 до 1,50 и общим средним значением 1.15. Восприимчивый класс ICR состоял из пяти человек со средним рейтингом заболевания в диапазоне 7,17–8,67 и общим средним значением 7,97 (таблица 2). Несколько человек (RK121, RK122 и RK173) проявили промежуточные реакции в диапазоне от 5,50 до 6,17 при заражении изолятом 6A и впоследствии были классифицированы как чувствительные. Однако из-за того, что средние показатели этих трех индивидуумов близки к среднему баллу по Комбар и резистентным к 6А индивидуумам, а также из-за того, что эти индивидуумы обладают высокой устойчивостью к изоляту 15А × 6А # 72 (VR2), эти очевидные промежуточные реакции могут неточно отражать классификация восприимчивых.
Разработка маркеров и построение карты с высоким разрешением
Использование синтении между ячменем и B. distachyon , ортологичные гены B. distachyon Bradi3g48220.1 и Bradi3g49360.1, соответствующие фланкирующим маркерам Spt1, и Rbah63b, соответственно , были идентифицированы с помощью поиска BLAST. Эта геномная область B. distachyon размером приблизительно 1,02 Mb между двумя ортологами содержит 149 аннотированных генов. B. distachyon. Гены , содержащие ортологи синтенного ячменя с интервалами приблизительно 60 т.п.н., были отобраны и использованы в поиске BLAST для идентификации высокодостоверных унигенов ячменя с поддержкой EST.Праймеры были сконструированы из 16 идентифицированных унигенов и использованы для получения генно-специфических ампликонов из сортов ячменя Rika и Kombar. Секвенирование по Сэнгеру было выполнено для выравнивания ампликонов и последовательностей, используемых для характеристики аллельных различий. Секвенирование 16 отобранных унигенов дало шесть устойчивых ампликонов, содержащих по крайней мере один SNP между родителями. Были сконструированы специфические для аллелей праймеры с 3′-концевыми нуклеотидами, специфичными для каждого родительского SNP, и их использовали в качестве маркеров STS для генотипирования ICR, чтобы насытить карту высокого разрешения маркерами в локусе Spt1 .Маркеры STS не могли быть получены из SNP в пределах двух ортологов ячменя, обозначенных как rpt-M8 и rpt-M14. Исходные пары праймеров использовали для получения ампликонов из каждого ICR, а затем секвенировали по Сэнгеру для генотипирования. Маркеры rpt-M4, rpt-M5 и rpt-M8 косегрегированы и картированы дистальнее локуса восприимчивости, rpt-M12 и rpt-M13 косегрегированы с Spt1 , а rpt-M14 картированы проксимальнее локуса восприимчивости. Недавно разработанные маркеры ограничили локус Spt1 до ∼0.43 см между дистальным фланкирующим маркером rpt-M8 и проксимальным фланкирующим маркером rpt-M14. Ограниченная область в ячмене соответствовала ~ 540 т.п.н. последовательности B. distachyon , которая содержала 69 аннотированных генов. Затем, используя вышеупомянутый процесс развития маркера посредством синтении, основное внимание было уделено дальнейшему насыщению региона. В общей сложности 17 ортологичных унигенов ячменя были идентифицированы из генов B. distachyon в этой области и нацелены на разработку маркеров, из которых секвенирование аллелей дополнительно позволило идентифицировать два ортолога, содержащие SNP.Вновь идентифицированные маркеры, rpt-M20 и rpt-M61, впоследствии были генотипированы посредством ПЦР-направленного генотипа-секвенирования целевых ампликонов из-за неспособности разработать эффективные маркеры STS. Кроме того, на основе того же ортолога, что и rpt-M61, был разработан надежный маркер indel, rpt-M62. Данные, полученные от чипа iSelect 9K для ячменя, также выявили маркер SNP, SCRI_RS_165041, который дополнительно ограничивает область. В настоящее время ген Spt1 ограничен маркерами rpt-M8 и SCRI_RS_165041 до ∼0.24 см регион. (Рисунок 1). Область ∼0,24 сМ, ограничивающая Spt1 в ячмене, соответствует ∼466 kb из последовательности B. distachyon , несущей 62 аннотированных гена, и, согласно прогнозам, будет содержать 49 генов ячменя с использованием данных привязки из базы данных генома ячменя и данных POPSEQ, а также as BLAST выполняет поиск генома ячменя, используя в качестве запроса гены B. distachyon . Попытки использовать эти гены для дальнейшего насыщения генетической карты выявили 15 ортологичных генов ячменя в этом регионе.Однако аллельное секвенирование интронов, 3 ‘UTR и кодирующих областей от Rika и Kombar этих выбранных ортологов определило, что эти гены не содержат полиморфизмов в нашей популяции (Рисунок 1)
Рисунок 1Генетическая и частичная физическая карта Spt1 Локус в синтении с Brachypodium distachyon . Последовательность Syntenous B. distachyon , карта высокого разрешения ячменя Rika × Kombar и сорт ячменя. Физическая карта Morex в локусе Spt1 . (A) Серые горизонтальные полосы представляют синтенозный B.distachyon хромосомы 3, отделяющей область гена Spt1 от ячменя. Нижняя полоса показывает относительное положение ортологичных последовательностей ячменя, используемых для разработки маркеров из аннотированных генов B. distachyon . Черные точки представляют позиции маркеров, а белые точки представляют собой фланговые маркеры Spt1 . Верхняя полоса представляет собой расширенное и более подробное представление ограниченной области Spt1 с аннотированными генами B. distachyon , показанными стрелками и помеченными выше с предполагаемыми функциями генов.Черные стрелки представляют гены, из которых ортологи ячменя использовались для генерации маркеров SNP (однонуклеотидный полиморфизм), а серые стрелки — это гены, которые не имели полиморфизмов в популяции Rika × Kombar, но были использованы для закрепления физических контигов на панели C. между стрелками указано количество дополнительных генов B. distachyon , присутствующих между каждым геном, обозначенным стрелкой. Масштаб 100 Кб показан вверху и внизу. (B) Рика × Комбарная хромосома 6H (Ch.6H) Spt1 карта высокого разрешения, полученная путем скрининга 2976 рекомбинантных F 2 гамет. Блоки содержат косегрегационные маркеры с белыми блоками, содержащими маркеры, которые разграничивают локус Spt1 . Числа между вертикальными линиями, соединенными с полями маркеров, указывают количество рекомбинантов между маркерами. Т и С в конце горизонтальной линии указывают направление теломер и центромеры соответственно. (C) Ячмень cv. Физическая карта Morex, показывающая относительные размеры суперконтигов (белые горизонтальные полосы) с приблизительным расположением генетических маркеров, сгенерированных из B.distachyon ортологичных генов ячменя. Черные кружки указывают положение маркеров, а белые кружки указывают фланговые маркеры области Spt1 . Положение генов ячменя, идентифицированных ортологами B. distachyon , из которых мы не смогли разработать маркеры, но которые могли закрепиться на физической карте ячменя, показано серыми стрелками. Проксимальный фланкирующий маркер SNP SCRI_RS_165041, идентифицированный по положению анализа ячменя 9k Illumina Infinium iSELECT, также обозначен белым кружком.Звездочкой обозначены два суперконтига, которые не были синтеническими, между B. distachyon и ячменем. Было обнаружено, что contigs_47325 и _47326 ячменя содержат ортологи ячменя аннотированных генов B. distachyon Bradi3g49130 и Bradi3g49140, которые в последовательности B. distachyon расположены между Bradi3g49110 и Bradi3g49160. Обозначения cv, Morex суперконтигов (IBGSC, 2012) приведены ниже в масштабе 1 Мб.
ПЦР-GBS секвенирование и вызов SNP
Секвенирование на платформе Ion Torrent PGM дало в общей сложности 569 040 прочтений со средней длиной 82 п.н.Однако из-за присутствия димеров праймеров только 184 174 чтения были специфичными для ампликона, что в среднем составляло ~ 5581 чтение на индивидуума, что достаточно для определения SNP для количества маркеров в этой панели. Ампликоны в библиотеке были представлены в разной степени из-за различий в эффективности ПЦР. Два маркера SNP из базы данных T3 (11_21216 и 11_20651) не были полиморфны между Rika и Kombar. Однако нам удалось идентифицировать полиморфизм между Rika и Kombar в ампликоне одного выбранного ортолога B.ген distachyon в области, обозначенной rpt-M32. Маркер SNP rpt-M32 картирован проксимальнее SCRI_RS_165041 и дистальнее rpt-M14 (рис. 1). Кроме того, два из четырех ранее идентифицированных маркеров STS (rpt-M4 и rpt-M8) имели достаточную репрезентативную последовательность для вызова SNP и подтверждали предыдущее генотипирование.
Скрининг библиотеки ВАС и построение физической карты
Скрининг библиотеки ВАС сорта ячменя Morex с маркерами rpt-M8, rpt-M13 и rpt-M12 идентифицировал четыре клона ВАС и был подтвержден с помощью ПЦР (таблица 3).Маркер rpt-M8 не амплифицировался ни на одной из идентифицированных ДНК-матриц ВАС. Оба маркера rpt-M12 и rpt-M13 были амплифицированы из ДНК-матриц ВАС 783N21, 302J12 и 082C13, что указывает на то, что эти клоны ВАС содержат MTP, в котором находятся оба маркера. Кроме того, с помощью поиска BLAST в базе данных HarvEST (Muñoz-Amatriaín et al. 2015) было подтверждено расположение маркеров rpt-M13 и rpt-M12 в клоне BAC 783N21, а также идентифицирован дополнительный BAC, 650G13, соответствующий к маркеру РПТ-М12.Все маркеры, использованные при генотипировании популяции картирования с высоким разрешением, также были подвергнуты поиску BLAST с использованием сервера HarvEST для идентификации BAC из Yu et al. (2000), в которой они находятся, что привело к идентификации соответствующих клонов ВАС для каждого маркера, за исключением rpt-M8 и rpt-M14 (Таблица 3). Кроме того, последовательности маркеров подвергали поиску методом BLAST с использованием сервера IPK Barley BLAST для идентификации соответствующих секвенированных клонов ВАС, cv. Morex WGS contigs и, если возможно, размещение на более крупных FPC.Маркеры rpt-M8, rpt-M20, rpt-M32 и rpt-M14 также использовали для скрининга cv. Библиотека Morex MTP BAC (INRA-CNRGV) для успешной идентификации соответствующих клонов BAC и позиции на FPC из-за невозможности привязать их к FPC с помощью методов поиска BLAST (таблица 3).
Таблица 3 Клоны бактериальной искусственной хромосомы (ВАС), идентифицированные по всем маркерам на карте высокого разрешения с соответствующими контигами последовательностей и контигами отпечатков пальцевОбсуждение
Предыдущие исследования устойчивости хозяина к NFNB в ячмене проводились с использованием различных массивов линий ячменя, а также а также грибковые изоляты из географически различных регионов мира.Результатом этих исследований была идентификация локусов устойчивости / восприимчивости, распределенных по геному ячменя (Ho et al. 1996; Steffenson et al. 1996; Raman et al. 2003; Cakir et al. 2003 ; Ма и др. 2004; Маннинен и др. 2006; Friesen и др. 2006a; Абу Камар и др. 2008; Сен-Пьер и др. 2010). Хотя доминантные локусы восприимчивости и / или устойчивости были нанесены на карту всех семи хромосом ячменя (обзор Liu et al. 2010), общий значимый QTL возле центромеры хромосомы 6H неоднократно был связан с устойчивостью и / или восприимчивостью к NFNB. Мы начали характеризовать этот важный локус, и вполне вероятно, что область QTL содержит несколько тесно связанных генов, которые взаимодействуют с предполагаемыми некротрофными эффекторами, лежащими в основе четырех основных QTL вирулентности, идентифицированных в P. teres f. teres Популяция патогенов 6A × 15A (Shjerve et al. 2014).Однако аллельный анализ генов-кандидатов, выявленных в этом исследовании, предполагает, что в основе этого главного локуса восприимчивости лежит единственный ген с дивергентными аллелями, которые придают доминантную восприимчивость калифорнийскому P. teres f. teres изоляты 15A и 6A (J. Richards и R. Brueggeman, неопубликованные данные).
Наличие множественных доминантных специфичностей восприимчивости (также известных как гены рецессивной устойчивости) в этом локусе (Abu Qamar et al. 2008; Liu et al. 2015) указывают на то, что репертуар эффекторных генов / генов вирулентности развился у патогена, чтобы воздействовать на сложную область рецепторов биотрофного иммунитета хозяина, чтобы вызвать NETS. Как видно в других патосистемах, специализирующихся на грибковых некротрофах, обозначение рецессивной резистентности, ранее данное этим генетическим взаимодействиям между хозяином и патогеном, вероятно, опосредовано функциональным продуктом гена доминантной восприимчивости, а не нефункциональным аллелем, и его следует более уместно описать как доминантную восприимчивость в инверсном гене. -для генной модели (Friesen et al. 2007). Подобно классической парадигме «ген за геном» (Flor 1956), продукт доминантного гена, продуцируемый хозяином, взаимодействует с продуктом доминантного гена патогена. Однако из-за некротрофного образа жизни этого специализированного патогена распознавание хозяином приводит к иммунному ответу PCD, который подрывается некротрофом, что приводит к совместимой реакции и болезни. Было замечено, что некоторые патогенные грибы некротрофных растений, такие как представители родов Pyrenophora и Parastagonospora , продуцируют эффекторы, которые распознаются одним геном-хозяином (Friesen et al. 2006b; Faris et al. 2010). Распознавание некротрофных эффекторов и последующая передача клеточных сигналов хозяином может происходить с помощью генов «устойчивости», кодирующих белки с сайтом связывания нуклеотидов (NBS), лейциновыми повторами (LRR) и / или доменами серин-треониновой протеинкиназы (STPK) (Lorang ). и др. 2007; Фарис и др. 2010 г.). Типичные механизмы ответа резистентности хозяина, такие как индукция запрограммированной гибели клеток, продукция активных форм кислорода (АФК) и повышающая регуляция генов PR, возникают после распознавания хозяином некротрофного эффектора (Lorang et al. 2007; Stergiopoulos et al. 2013; Лю и др. 2015). Хотя эти ответы обеспечивают эффективную защиту от биотрофных патогенов, некротрофы, которые выживают во враждебной среде хозяина, создаваемой в качестве барьера для колонизации, также намеренно вызывают иммунные ответы PCD для получения питательных веществ из образовавшейся мертвой ткани хозяина. Однако, в отличие от биотрофной устойчивости хозяина этими путями, которые обычно обеспечивают качественную доминантную резистентность, эти обратные взаимодействия ген-ген, по-видимому, носят количественный характер (Friesen et al. 2008).
Посредством картирования с высоким разрешением популяции Rika × Kombar F 2 с использованием маркеров, полученных из ортологичных генов B. distachyon в ранее выделенной области rpt.k / rpt.r , мы определили предполагаемую доминантную восприимчивость локус, обозначенный Spt1 , в область ~ 0,24 сМ вблизи центромеры хромосомы 6H ячменя. Это картирование с высоким разрешением значительно уточнило ранее ограниченную область ∼3,3 сМ и облегчило построение физической карты с использованием новой последовательности генома ячменя.Использование различных недавно доступных геномных ресурсов ячменя, которые быстро уточняются и обновляются, ускорило построение генетической карты в физическую и идентификацию гена кандидата Spt1 . Хотя физическая карта области Spt1 не является полностью смежной, эти инструменты позволили расположить физические последовательности в корреляции с нашей картой высокого разрешения, а также идентифицировать фланкирующие клоны ВАС, которые будут использоваться для дальнейшей разработки маркеров и насыщенность карты.Из-за идентификации контигов ячменя, содержащих синтенные гены B. distachyon , мы ожидаем, что мы захватили большую часть физического региона Spt1 и покажем, что он был ограничен примерно 9,5 МБ генома ячменя, что прогнозируется. содержать 49 генов.
Мы предполагаем, что ген / гены, лежащие в основе локуса Spt1 в охарактеризованной области, придают доминантную восприимчивость к изолятам 6A и 15A NFNB за счет распознавания некротрофных эффекторов, продуцируемых патогеном, и последующей индукции путей PCD в организме хозяина.Недавно было показано, что P. teres f. teres продуцирует некротрофные эффекторы, которые вызывают вышеупомянутые ответы хозяина (Liu et al. 2015). Используя межклеточные промывочные жидкости из ткани листьев ячменя, инокулированной P. teres f. teres изолят 0-1, некроз был индуцирован у восприимчивого сорта Гектор, и чувствительность впоследствии была картирована в центромерной области хромосомы 6H. Анализ сегрегации показал, что единственный локус на 6H, вероятно, содержит ген, придающий чувствительность к эффектору, продуцируемому P.teres f. teres изолят 0-1 (Liu et al. 2015). Кроме того, четыре локуса вирулентности ( VR1 , VR2 , VK1 и VK2 ) были картированы в P. teres f. teres. картируют популяцию, полученную из скрещивания изолятов 6A и 15A, идентифицируя два основных QTL вирулентности для каждого изолята в разных регионах P. teres f. терес генома. Изоляты потомства, содержащие одиночные локусы вирулентности, были идентифицированы для VK1 , VK2 и VR2 , по существу, Менделируя основной QTL вирулентности из 15A и один из 6A путем выделения специфических некротрофных взаимодействий эффектор-хозяин.Эти отдельные изоляты потомства QTL были инокулированы в критические рекомбинанты 6H для картирования генов чувствительности, которые, как мы ожидаем, будут кодировать белки, подобные рецепторам иммунитета, которые регулируют чувствительность, опосредованную этими уникальными эффекторами. Восприимчивость к изолятам-потомкам одной основной вирулентности VR2 , VK1 и VK2 , картированным в одну и ту же область на 6H и, что интересно, все картированы в локусе Spt1 . Это указывает на то, что вирулентность, обеспечиваемая изолятом 15А, может регулироваться распознаванием двух уникальных некротрофных эффекторов двумя тесно связанными генами.Альтернативно, единственный ген в локусе Spt1 может воспринимать оба некротрофных эффектора, продуцируемые изолятом 15A. Предполагаемый некротрофический эффектор, продуцируемый изолятом потомства, несущим VR2 , придающий вирулентность изоляту 6A, может также прямо или косвенно взаимодействовать с доминантным фактором восприимчивости в локусе Spt1 , который может быть или не быть аллелем гена, придающего восприимчивость. изолировать 15А. Альтернативно, несколько эффекторных мишеней в пределах ∼0.24 cM Spt1 генетический интервал может существовать на «острове восприимчивости», каждый из которых распознает уникальный некротрофный эффектор и вызывает PCD. Мы определили генетическую область примерно на 9,5 МБ физической последовательности ячменя, которая на основе POPSEQ якорения унигенов ячменя и поисков BLAST ортологичных генов B. distachyon внутри области против последовательности генома ячменя, содержит прогнозируемые 39 с высокой степенью достоверности. гены. На основании аннотации функциональных доменов шесть генов кодируют предсказанные рецептороподобные белки иммунитета.Таким образом, эти гены являются генами-кандидатами Spt1 и далее анализируются с помощью анализа аллелей и ассоциативного картирования с высоким разрешением.
Патосистема ячменя-NFNB оказывается генетически сложной, состоящей из доминантных, рецессивных и неполных резистентностей, которые сгруппированы в локусе 6H. Эта сложность 6H усложняет попытки включить эффективную устойчивость в элитные линии ячменя. Клонирование Spt1 , а также эффекторов, которые взаимодействуют с ними прямо или косвенно, позволит лучше узнать и понять молекулярные механизмы, лежащие в основе этой патосистемы, а также то, как специализированные грибковые патогены-некротрофы эволюционировали, чтобы использовать сложный иммунитет. система хозяина, чтобы вызвать полезные ответы PCD.Выделение отдельных генов устойчивости / восприимчивости, вовлеченных в эту патосистему, заполнит пробелы в понимании молекулярных основ взаимодействий некротрофного специалиста и хозяина и позволит исследователям эффективно использовать идентифицированные источники устойчивости. Эта работа значительно продвигает усилия по позиционному клонированию и функциональному анализу генов-хозяев, вовлеченных в эту сложную патосистему.
Благодарности
Авторы хотели бы поблагодарить Даниэль Холмс за техническую помощь Служба сельскохозяйственных исследований Министерства сельского хозяйства США (USDA-ARS), Отдел исследования зерновых культур, Фарго, Северная Дакота.Этот проект был поддержан Сельскохозяйственным проектом по координации конкурсных грантов Национальной исследовательской инициативы (CAP), проектом 2011-68002-30029 Национального института продовольствия и сельского хозяйства США (NIFA), Национальным научным фондом (NSF), экспериментальной программой Северной Дакоты по стимулированию Конкурентные исследования (NDEPSCoR), грант Track 1 11A-1355466 и номер награды проекта NSF CAREER 1253987.
- Получено 11 марта 2016 г.
- Принято 14 апреля 2016 г.