
как работают наши вкусовые рецепторы
Делимся переводом статьи про анатомию вкуса. Она поможет понять, как формируется вкусовое впечатление и почему в оценке кофе важен не только вкус, но и тело, запах и температура.
Вкусовые сосочки: из чего состоит поверхность языкаЕсли внимательно посмотреть на язык, мы увидим, что он как будто покрыт маленькими «пупырышками». Кажется, что это и есть наши вкусовые рецепторы. Но на самом деле это вкусовые сосочки. Рецепторы намного меньше: в одном вкусовом сосочке — от 3 до 100 рецепторов.
Вкусовые сосочки языка
Обратите внимание, что некоторые сосочки выглядят иначе, чем другие. Это потому что у нас на языке четыре типа сосочков. В центре языка — множество маленьких, тонких сосочков. Они называются нитевидными и не содержат вкусовых рецепторов. Спереди и по бокам языка — маленькие круглые сосочки, похожие на крапинки.
Увидеть другие два типа гораздо сложнее. Посмотрите на самую заднюю часть языка — туда, где расположены миндалины. Возможно, вам удастся разглядеть несколько выступов по бокам. Это листовидные сосочки. А ближе к небному язычку расположены большие, круглые вкусовые сосочки. Они называются желобовидными. Каждый из этих двух типов содержит более 100 вкусовых рецепторов.
Сладкий, горький, солёный. Правда ли, что разные части языка отвечают за разные вкусы?Глядя на расположение сосочков на языке, мы вспоминаем уроки биологии, на которых изучали карту языка. Она показывает области, которые отвечают за распознавание разных вкусов — соленого, сладкого, кислого, горького и острого. Но не все так просто: карта языка — это миф! Каждый вкусовой рецептор, независимо от расположения, может распознавать все пять вкусов.
Карта языка — это миф: каждый вкусовой рецептор распознает все пять вкусов
Давайте поближе рассмотрим вкусовой рецептор.
Каждая вкусовая клетка имеет вкусовой волосок. Когда он соприкасается с молекулой пищи, клетка посылает мозгу импульс: обнаружен вкус! Импульс проходит через черепной нерв, который соединяет вкусовые рецепторы с мозгом.
Каждый вкусовой рецептор содержит 30–50 вкусовых клеток. Это означает, что ваш мозг одновременно обрабатывает тысячи сигналов от вкусовых рецепторов. Он берет информацию, которую посылают рецепторы, и сравнивает с воспоминаниями о вкусах, которые вы когда-либо ощущали. Именно так мозг распознает, что именно вы едите.
Существует всего пять вкусов — сладкий, соленый, кислый, горький и умами. Все остальное ваши вкусовые рецепторы попросту игнорируют. Но мы знаем, что вкусовые впечатления нельзя уместить в пять определений. И в этом случае пора вспомнить про запах и ощущения во рту.
И в этом случае пора вспомнить про запах и ощущения во рту.
Когда мы говорим «вкусно», наш мозг на самом деле оценивает не только вкус, но и множество других факторов. Текстура, влажность, температура, жирность — это лишь часть того тактильного опыта, который мы получаем во время жевания или глотания. Вы не станете есть отсыревшую гранолу, пить теплую газировку или угощать друзей засохшим печеньем. Даже если вкус блюд остался тем же, общее впечатление будет негативным.
Как запах влияет на восприятие вкусаВо время жевания и глотания вы ощущаете запахи даже той пищи, которая уже у вас во рту. Все они поступают в носовую полость, где расположены обонятельные клетки. Эти клетки работают так же, как и вкусовые рецепторы — но отвечают за восприятие запаха, а не вкуса. У них тоже есть крошечные волоски, которые при контакте с запахом посылают мозгу сигнал через обонятельный нерв.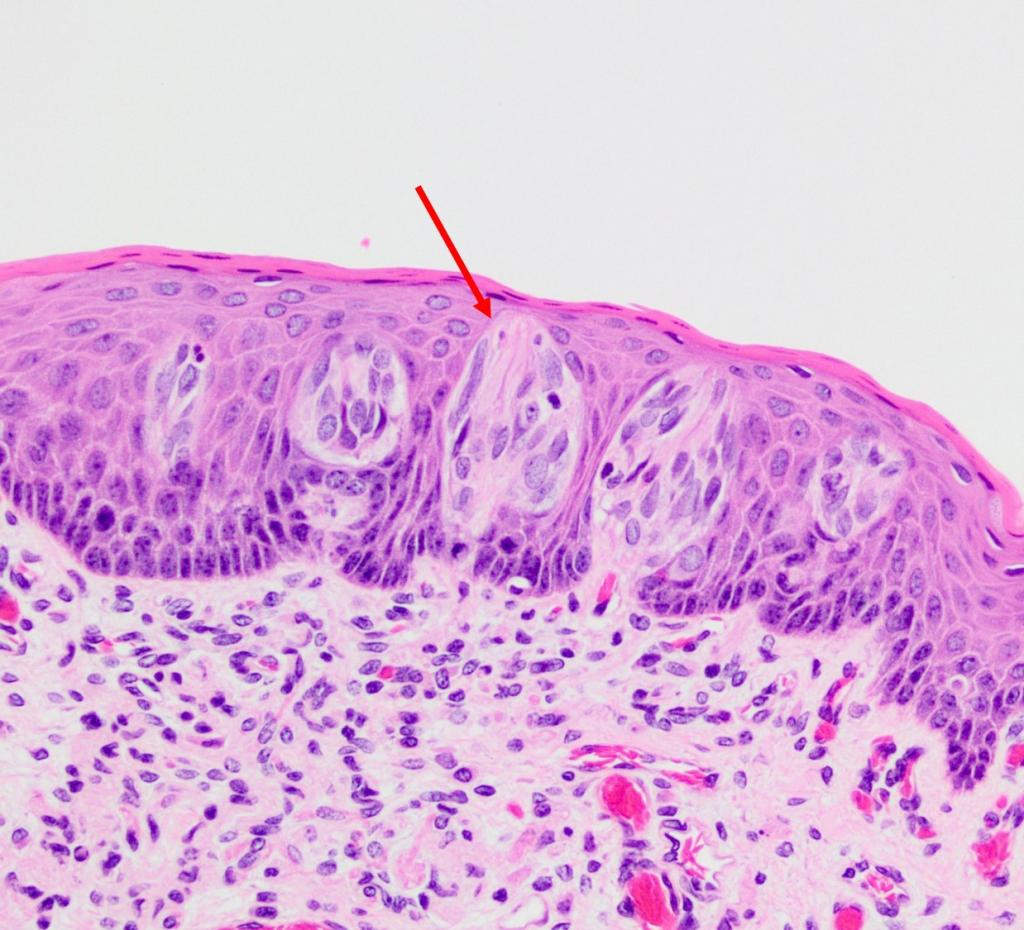
Взаимодействие обонятельных клеток с запахами
В отличие от вкусовых, обонятельные клетки могут различать тысячи разных ароматов — от мясного до фруктового. Получается, что практически каждый компонент нашего вкусового впечатления, кроме пяти известных вкусов, — это на самом деле запах. Поэтому он так важен в восприятии вкуса. Это также объясняет, почему при заложенном носе пища кажется безвкусной.
Нам кажется, что во время приема пищи активна только наша пищеварительная система, но это не так. Наш мозг проделывает огромную работу. А работает он в тандеме с вкусовыми и обонятельными рецепторами — именно они помогают нам распознавать вкусы. Благодаря сложной анатомии мы помним любимые лакомства из детства и можем наслаждаться блюдами в кафе.
Оригинал: http://www.decodingdelicious.com/the-anatomy-of-flavor/
| | Поместите стрелку мыши или нажмите пальцем на фотографию и Вы сможете увидеть ее без обозначений (при медленной загрузке — не убирайте стрелку мыши или палец |
| НИТЕВИДНЫЕ СОСОЧКИ ЯЗЫКА Окраска гематоксилин-эозином 1 — эпителий, покрывающий сосочек | |
| ЛИСТОВИДНЫЕ СОСОЧКИ ЯЗЫКА Окраска гематоксилин-эозином 1 — эпителий, покрывающий сосочек | |
| ЖЕЛОБОВАТЫЙ СОСОЧЕК ЯЗЫКА Окраска гематоксилин-эозином 1 — эпителий, покрывающий сосочек | |
| ШЛИФ ЗУБА Неокрашенный препарат 1 — эмаль | |
| РАЗВИТИЕ ЗУБА — ЭМАЛЕВЫЙ ОРГАН, ЗУБНОЙ
СОСОЧЕК, ЗУБНОЙ МЕШОЧЕК Окраска гематоксилин-эозином 1, 3, 4 — эмалевый орган 2 — зубной сосочек 3 — наружные клетки эмалевого органа 4 — промежуточные клетки эмалевого органа 5 — зубной мешочек 6 — эпителий полости рта | |
| РАЗВИТИЕ ЗУБА — ОБРАЗОВАНИЕ ТКАНЕЙ ЗУБА Окраска гематоксилин-эозином 1 — эмалеобласты | |
| РАЗВИТИЕ ЗУБА — ОБРАЗОВАНИЕ ТКАНЕЙ ЗУБА Окраска гематоксилин-эозином 1 — эмалеобласты обозначена пунктирной линией |
Система органов пищеварения :: Вкусовые сосочки
Система органов пищеварения
Вкусовые сосочки
Вкусовые сосочки по форме и топографии подразделяются на грибовидные, валиковидные и листовидные.
Валиковидные сосочки (p. vallatae) довольно крупные, округлой или овальной формы. Вкусовые луковицы находятся на боковых стенках ровиков (см. рис. 7, 8 Г).
Рис. 8. Сосочки языка лошади (по Элленбергеру): А – нитевидный: |
На дне ровиков открываются протоки концевых белковых желез языка, располагаются они в области корня языка. У жвачных количество их достигает 17 пар (коровы), а у диких жвачных и более (олени – до 37). У собаки и свиньи их мало – 1-3 пары. У лошади их всего одна пара, но довольно крупных размеров. Валиковидные сосочки воспринимают горький вкус.
У жвачных количество их достигает 17 пар (коровы), а у диких жвачных и более (олени – до 37). У собаки и свиньи их мало – 1-3 пары. У лошади их всего одна пара, но довольно крупных размеров. Валиковидные сосочки воспринимают горький вкус.
Грибовидные сосочки (p. fungiformes) располагаются на дорсальной поверхности тела и верхушки языка (см. рис. 7, 8 Б). Наибольшее количество их на кончике и боковых краях тела. Диаметр до 1 мм. Вкусовые луковицы расположены по всей поверхности стенки сосочка. Они имеют продолговато-овальную форму. Периферические клетки вкусовой луковицы светлые, центральные имеют темную цитоплазму и овальное ядро. Грибовидные сосочки воспринимают раздражающий вкус – соленое и кислое.
Листовидные сосочки (p. foliatae) имеются по бокам корня языка, образованы складками слизистой оболочки языка (см. рис. 7, 8 В). Вкусовые луковицы лежат в стенке ровиков, на дне которых открываются протоки концевых белковых желез языка. Листовидных сосочков нет у крупного рогатого скота, овец и кошек. Листовидные сосочки воспринимают сладкий вкус.
Листовидные сосочки воспринимают сладкий вкус.
как язык сообщает мозгу вкус пищи
Ореховый привкус выдержанного сыра, ягодные нотки вин, цветочное послевкусие темного шоколада — восхитительные вкусы постоянно бомбардируют наши языки. Мы все любим вкусно покушать, но задумывались ли вы о том, как язык переносит эти божественные вкусы с тарелки в мозг?
Язык, который отвечает за вкусовую систему организма, представляет собой кусок мышечной плоти, покрытый слизистой оболочкой. Человеческому глазу наши языки кажутся усеянными крошечными бугорками, называемыми сосочками. Их часто ошибочно принимают за сами вкусовые рецепторы, однако на самом деле эти сосочки содержат группы вкусовых рецепторов.
Язык усеян тремя типами сосочков, воспринимающих вкус. Грибовидные сосочки, сосредоточенные в основном на кончике и боковых сторонах языка, обычно содержат один вкусовой рецептор на своих грибовидных кончиках. Листовидные сосочки, расположенные в красноватых складках по бокам языка, содержат множество вкусовых рецепторов, организованных вокруг этих щелей. Более крупные куполообразные папиллярные сосочки расположены ближе к задней части языка, и, подобно листовидным сосочкам, они могут вместить до 250 вкусовых рецепторов каждый.
Листовидные сосочки, расположенные в красноватых складках по бокам языка, содержат множество вкусовых рецепторов, организованных вокруг этих щелей. Более крупные куполообразные папиллярные сосочки расположены ближе к задней части языка, и, подобно листовидным сосочкам, они могут вместить до 250 вкусовых рецепторов каждый.
Четвертый тип — нитевидные сосочки — самый маленький и самый многочисленный на языке. Этот тип содержит тонкие волоски, которые соединяются с нервами, связанными с осязанием, позволяя нам чувствовать текстуру того, что мы едим, но при этом они не содержат никаких вкусовых рецепторов.
В то время как эти различные типы сосочков различаются по структуре, они обычно группируются вокруг щелей, в которые попадают разрушенные пищевые химические вещества, высвобождающиеся при жевании. Эти щели также содержат железы, которые выделяют слюну.
Вкусовые рецепторы тоже представляют собой сложные небольшие бугорки. Каждый содержит от 50 до 150 специальных клеток, которые располагаются как зубчики в луковице чеснока и реагируют на определенные химические соединения. Общее количество вкусовых рецепторов от человека к человеку разнится, и может составлять от 4000 до 10000 штук.
Общее количество вкусовых рецепторов от человека к человеку разнится, и может составлять от 4000 до 10000 штук.
Каждый вкусовой рецептор специализируется на обнаружении одного из пяти типов вкуса: сладкого, кислого, горького, соленого или умами. Да, последний вкус известен нам мало — он есть в соевом соусе и в сыре пармезан, также его дает усилитель вкуса глутамат натрия. Зато в Китае и Японии этот вкус высокобелковых веществ очень распространен.
Вопреки тому, что вы, возможно, учили в школе, на языке нет областей, специализирующихся на каком-то определенном вкусе — вместо этого каждый вкусовой сосочек имеет все пять вкусовых рецепторов, встроенных в него. Каждый рецептор имеет небольшие волоскоподобные клетки, называемые микроворсинками, которые связываются с определенными химическими соединениями, соответствующими типу вкуса, на котором он специализируется.
Увы — ученые все еще выясняют, как именно химические вещества взаимодействуют с рецепторами. Глубже в плоти языка они прикрепляются к нервам, которые соединяются с остальной нервной системой и передают информацию о вкусе в мозг.
Итак, после такого достаточно длинного введения можно перейти к подробностям. Когда вы едите, ваш рот разделывает пищу на ее химические строительные блоки. Ваши зубы физически измельчают и разделяют крупные компоненты еды, а слюна содержит различные ферменты, которые разбивают эти крошки на молекулы:
Как только эти пищевые химические вещества начинают плавать вокруг языка, часть из них попадает в пространство между сосочками, забивается в щели и достигает вкусовых рецепторов. Микроворсинки последних взаимодействуют только с определенными химическими веществами, которые соответствуют их вкусовой специализации.
Сахар запускает сладкие рецепторы, кислоты запускают кислые рецепторы, минералы, такие как хлорид натрия и магний, запускают солевые рецепторы, группа из около 35 белков запускает горькие рецепторы, а глутаминовая и аспарагиновая кислоты запускают умами:
Как только микроворсинки связываются с определенными химическими веществами, рецепторы активируют свои индивидуальные нервы. И в итоге они посылают целую бурю электрических сигналов в мозг, чтобы сказать нашей голове о том, что эти вкусы присутствуют на языке. По этой причине вкус первого кусочка нам кажется максимально ярким и полным. В конце концов, когда рецепторные клетки привыкают к вкусу, в мозг начинают поступать более упорядоченные сигналы, и последний правильно их интерпретирует — в итоге вкус еды приглушается:
И в итоге они посылают целую бурю электрических сигналов в мозг, чтобы сказать нашей голове о том, что эти вкусы присутствуют на языке. По этой причине вкус первого кусочка нам кажется максимально ярким и полным. В конце концов, когда рецепторные клетки привыкают к вкусу, в мозг начинают поступать более упорядоченные сигналы, и последний правильно их интерпретирует — в итоге вкус еды приглушается:
Нос тоже в деле: еда имеет и свои запахи, которые при измельчении во рту только усиливаются. В итоге они проходят через заднюю часть рта и попадают в носовую полость. Здесь обонятельные сенсоры обнаруживают более сложные вкусовые профили, помимо пяти закодированных вкусовыми рецепторами на языке, и посылают сигналы о них в мозг:
Информация от вкусовых рецепторов вместе с аналогичными сигналами от других чувствительных нервов, отвечающих за боль, температуру и осязание, достигает нижнего отдела мозгового ствола. Оттуда данные о вкусе переходят уже в сознательные отделы головного мозга:
Ну и, пожалуй, самая приятная часть: как только вкусовые сигналы достигают областей сенсорного восприятия в мозге, они объединяются с сигналами обоняния, чтобы сообщить голове, что находится во рту. И если мы воспримем общий вкус как приятный, то мозг вызовет увеличение производства слюны и желудочного сока, способствуя пищеварению и делая еду еще более аппетитной:
И если мы воспримем общий вкус как приятный, то мозг вызовет увеличение производства слюны и желудочного сока, способствуя пищеварению и делая еду еще более аппетитной:
Как видите, распознавание вкуса — дело непростое, за него отвечают тысячи нейронов и вкусовых рецепторов, и участвует в этом даже нос. Так что такой, казалось бы, достаточно обыденный процесс — поглощение пищи — своего рода биохимическое чудо.
iGuides в Telegram — t.me/igmedia
iGuides в Яндекс.Дзен — zen.yandex.ru/iguides.ru
Язык как зеркало здоровья | Интернет-издание «Новости медицины и фармации»
Все внутренние изменения человеческого тела: состояния недостатка или избыточности внутренних органов,
любые состояния недавней или развившейся болезни, состояния полноты или недостатка жидкостей организма,
состояния силы или упадка Ци и Крови — могут быть объективно отражены характерными изменениями языка.
Доктор Ze-Lin Chen, Китай
В статье представлены данные литературы и собственные клинические наблюдения за состоянием слизистой оболочки полости рта у детей с различной патологией. Оценка состояния языка важна при описании объективного статуса пациента, особенно в динамике наблюдения. Ее отражение в медицинской документации является простым и важным скрининговым показателем для составления алгоритма диагностического поиска, а также дополнительным критерием эффективности лечения, реабилитации и профилактики различных заболеваний.
С древних времен врачи пытались оценить состояние здоровья пациента по его внешним признакам, так как уже знали о единстве организма и взаимосвязи всех органов и систем, а также о том, что нарушение функции одного из них приводит к изменениям в другом.
Несмотря на то, что в настоящее время существует огромный арсенал различных лабораторных и инструментальных диагностических методов, основанных на современных технологиях, по-прежнему важным остается оценка врачом объективного статуса пациента. Недаром народная мудрость гласит, что перед любым современным аппаратом «надо снимать шляпу, а голову оставлять на плечах».
Недаром народная мудрость гласит, что перед любым современным аппаратом «надо снимать шляпу, а голову оставлять на плечах».
Согласно постулатам Аюрведы (древнеиндийской науки о здоровье) и китайской медицины, изменение пульса на лучевой артерии и состояния кожи и слизистых оболочек, в том числе языка, несет в себе информацию о наличии ряда заболеваний [1, 2]. Еще Гиппократ в описаниях болезней придавал большое значение изменениям на слизистой оболочке языка. Так, при плевропневмонии он указывал на зеленоватый цвет в верхней его части, при желтухе — желтый цвет и толстые черные подъязычные вены [3]. В настоящее время оценка состояния языка остается важной при осмотре больного в Тибете, Индии и Китае.
Язык — это мышечный орган, участвующий в механической обработке пищи, акте глотания, восприятии вкуса, формировании речи. Слизистая оболочка языка достаточно устойчива к действию различных механических, химических и термических факторов во время приема пищи и воды. Ей свойственны высокая регенераторная способность (обновление каждые три дня), а также относительная устойчивость к внедрению инфекции. В поверхностном слое слизистой оболочки располагаются сосочки, являющиеся органами вкусовой и тактильной чувствительности. Сосочки разделяются по форме, размеру и функции на четыре вида: нитевидные, желобоватые, грибовидные и листовидные. Нитевидные сосочки наиболее многочисленны (до 500 на 1 см2), покрыты ороговевающим эпителием, играют роль органов осязания и выполняют механическую функцию. Грибовидные сосочки в количестве от 40–50 до 90 на 1 см2 располагаются среди нитевидных, их больше на кончике языка и в средней части спинки. Эпителий, покрывающий эти сосочки, не ороговевает. В грибовидных сосочках заложены терморецепторы и так называемые вкусовые почки, в которых сконцентрированы 50–100 клеток вкусовых рецепторов. Новорожденные обычно имеют больше вкусовых почек, чем взрослые, у которых часть из них с возрастом дегенерирует и исчезает. Желобоватые сосочки в количестве от 7 до 12 также являются органами вкуса, располагаются на границе между корнем и телом языка, не выступают над поверхностью слизистой оболочки.
В поверхностном слое слизистой оболочки располагаются сосочки, являющиеся органами вкусовой и тактильной чувствительности. Сосочки разделяются по форме, размеру и функции на четыре вида: нитевидные, желобоватые, грибовидные и листовидные. Нитевидные сосочки наиболее многочисленны (до 500 на 1 см2), покрыты ороговевающим эпителием, играют роль органов осязания и выполняют механическую функцию. Грибовидные сосочки в количестве от 40–50 до 90 на 1 см2 располагаются среди нитевидных, их больше на кончике языка и в средней части спинки. Эпителий, покрывающий эти сосочки, не ороговевает. В грибовидных сосочках заложены терморецепторы и так называемые вкусовые почки, в которых сконцентрированы 50–100 клеток вкусовых рецепторов. Новорожденные обычно имеют больше вкусовых почек, чем взрослые, у которых часть из них с возрастом дегенерирует и исчезает. Желобоватые сосочки в количестве от 7 до 12 также являются органами вкуса, располагаются на границе между корнем и телом языка, не выступают над поверхностью слизистой оболочки. Валик слизистой оболочки, окружающий каждый сосочек, отделяется от него глубокой бороздой, куда открываются выводные протоки мелких серозных слюнных желез Эбнера. Листовидные сосочки числом 15–20 состоят из вертикальных складок, которые располагаются в основании боковой поверхности языка. В их эпителии тоже находятся вкусовые луковицы. Различные участки поверхности языка обладают неодинаковой чувствительностью к вкусовым раздражителям. Так, к горькому вкусу более чувствителен корень языка, к кислому — боковые поверхности, к соленому — боковые поверхности и кончик, к сладкому — кончик языка [4, 5].
Валик слизистой оболочки, окружающий каждый сосочек, отделяется от него глубокой бороздой, куда открываются выводные протоки мелких серозных слюнных желез Эбнера. Листовидные сосочки числом 15–20 состоят из вертикальных складок, которые располагаются в основании боковой поверхности языка. В их эпителии тоже находятся вкусовые луковицы. Различные участки поверхности языка обладают неодинаковой чувствительностью к вкусовым раздражителям. Так, к горькому вкусу более чувствителен корень языка, к кислому — боковые поверхности, к соленому — боковые поверхности и кончик, к сладкому — кончик языка [4, 5].
Согласно канонам китайской медицины, все внутренние органы человека имеют зону проекции на определенные участки языка (рис. 1). Так, кончик несет информацию о состоянии сердца; зона перед кончиком языка отражает состояние легких; центральная часть языка — желудка, поджелудочной железы; средняя треть, латеральнее срединной борозды — селезенки, печени и желчного пузыря; задняя часть — почек; корень языка — кишечника [4, 5]. Следовательно, по внешнему виду языка можно судить о состоянии внутренних органов пациента. Для этого необходимо определить размер, структуру поверхности, цвет и влажность языка [2].
Следовательно, по внешнему виду языка можно судить о состоянии внутренних органов пациента. Для этого необходимо определить размер, структуру поверхности, цвет и влажность языка [2].
У здорового человека слизистая оболочка языка бледно-розового цвета, без налета, с ровной складкой посередине, хорошо выраженными сосочками. В жаркое время года сосочки языка имеют красноватый оттенок и немного увеличены. У здоровых детей раннего возраста слизистая оболочка языка розового цвета, выражены V-образно стоящие валикообразные сосочки. Листовидные сосочки на боковых поверхностях языка появляются только к 6–7 годам [6].
На состояние слизистой оболочки языка оказывают влияние состав пищи, травмы (механические, термические, химические), токсико-аллергические воздействия (стоматологические материалы, лекарственные вещества и др.) [6, 7]. Так, острая пища и пряности (маринады, кайенский перец, карри и др.) оказывают раздражающее действие и на короткое время придают языку красный цвет, употребление крепкого кофе — коричневый, свеклы — фиолетовый. Курение может стать причиной желтого налета, прием антибиотиков — привести к частичному исчезновению участков эпителия на поверхности языка.
Курение может стать причиной желтого налета, прием антибиотиков — привести к частичному исчезновению участков эпителия на поверхности языка.
При инфекционно-воспалительных заболеваниях язык становится гиперемированным, при хронических заболеваниях и анемии — бледным, при дискинезии желчевыводящих путей, холецистите, вирусном гепатите — желтым. При заболеваниях легких и сердечно-сосудистой системы язык приобретает цианотичный оттенок.
Анализируя состояние языка, важно отметить качество и количество имеющегося налета. У здорового человека, допустим, тонкий налет белого цвета, легко удаляющийся зубной щеткой или щеточкой для чистки языка. Белый, плотный налет в зависимости от зоны расположения на языке является признаком воспалительного процесса какого-либо органа, а также может быть проявлением эндогенной интоксикации (при запорах, наличии в организме токсических и условно-токсических макро- и микроэлементов, почечной и печеночной недостаточности и др.) (рис. 2). Налет коричневого цвета может появляться у больных с заболеваниями легких и кишечника, желтого цвета — при застойных явлениях в желчном пузыре.
Вертикальные бороздки по краям языка отмечаются при различных заболеваниях желудочно-кишечного тракта (рис. 3).
Неровная продольная борозда посередине языка может наблюдаться у пациентов с нарушением осанки. При этом ее расположение в передней трети языка свидетельствует об искривлении шейного, средней трети — грудного, задней трети — поясничного отдела позвоночника (рис. 4).
Утолщение языка возникает при различной патологии печени. Отпечатки зубов на боковых поверхностях языка появляются у больных с хроническим энтероколитом, диспепсией. Сухой язык, многочисленные трещины наблюдаются при лихорадке, диарее, диабете, анемии. Тремор языка может быть проявлением неврастенического синдрома, вегетоневроза, тиреотоксикоза [2, 5–8].
Особое внимание необходимо уделять наличию так называемого географического языка, что клинически проявляется участками гиперемии, чередующимися с участками серого цвета до 0,5 см в диаметре, которые отграничены друг от друга желтовато-серыми валиками, состоящими из утолщенных нитевидных сосочков (рис.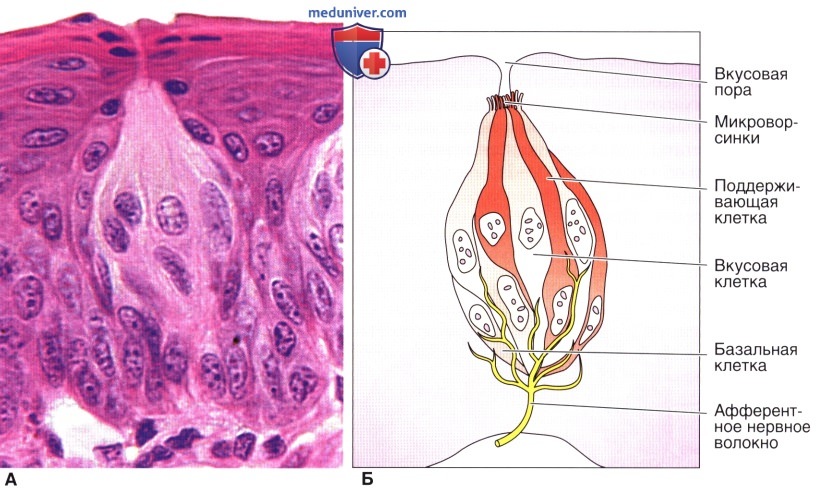 5). В течение нескольких дней эти участки могут изменять свою форму и локализацию. Сливаясь друг с другом, они создают на языке рисунок, напоминающий географическую карту. Впервые это состояние описал французский врач Rayer в 1831 г. Такая десквамация эпителия слизистой оболочки языка характерна для больных с атопическим дерматитом, глистной инвазией, гастроэзофагеальной рефлюксной болезнью, дискинезией желчевыводящих путей, гастритом со сниженной секреторной функцией, вирусным гепатитом [2, 6, 8].
5). В течение нескольких дней эти участки могут изменять свою форму и локализацию. Сливаясь друг с другом, они создают на языке рисунок, напоминающий географическую карту. Впервые это состояние описал французский врач Rayer в 1831 г. Такая десквамация эпителия слизистой оболочки языка характерна для больных с атопическим дерматитом, глистной инвазией, гастроэзофагеальной рефлюксной болезнью, дискинезией желчевыводящих путей, гастритом со сниженной секреторной функцией, вирусным гепатитом [2, 6, 8].
Наиболее часто изменениями в полости рта сопровождаются заболевания желудочно-кишечного тракта. Это объясняется общностью морфологического строения слизистой оболочки рта и пищеварительного тракта [9]. Так, по данным Е.Г. Романенко, обследовавшей 162 ребенка с хроническим гастродуоденитом, у 76,3 % из них отмечался гиперкератоз нитевидных сосочков, у 69 % — гиперемия и отечность языка, у 64 % обследуемых — гипертрофия грибовидных и листовидных сосочков. У детей с повышенной кислотообразующей функцией желудка наблюдалось усиление слюноотделения, что обусловлено параллелизмом в работе слюнных желез и желез фундального отдела, вырабатывающих слизь [10]. Часто у пациентов с гиперсаливацией отмечался отек языка с отпечатками зубов на боковых поверхностях (рис. 6).
Часто у пациентов с гиперсаливацией отмечался отек языка с отпечатками зубов на боковых поверхностях (рис. 6).
Для гастроэзофагеальной рефлюксной болезни патогномоничны множественная деминерализация твердых тканей зубов, наличие географического языка и/или налет белого цвета, не имеющий четкой локализации. При хроническом гастродуодените без нарушения моторной функции налет расположен равномерно [10]. При язвенной болезни желудка и двенадцатиперстной кишки язык отечный, покрыт тяжело снимающимся налетом, иногда — клейкой слизью [8]. При хронических заболеваниях печени на языке часто появляются геморрагии [8, 9, 11]. Для острого панкреатита характерна сухость языка, увеличение количества нитевидных и гиперплазия грибовидных сосочков, при хроническом панкреатите могут наблюдаться атрофические процессы [8]. При заболеваниях кишечника в полости рта могут появляться множественные, резко болезненные эрозии (афты), налет серо-желтого или коричневого цвета [7, 8, 11].
Изменения слизистой оболочки полости рта имеют место и при эндокринных заболеваниях. Так, при гипофункции щитовидной железы отмечается сухость языка и губ, при ее гиперфункции — тремор, снижение вкуса [1, 11]. Утолщение языка и образование складок на нем (за счет увеличения количества нитевидных сосочков) в совокупности с другими симптомами могут иметь значение в ранней диагностике нарушения функции гипофиза [1]. При хронической недостаточности коры надпочечников на слизистой оболочке полости рта и боковых поверхностях языка появляются коричневые пятна различной формы без признаков воспаления. При гиперфункции коры надпочечников в полости рта могут быть выявлены афты, участки гиперкератоза, кандидомикоза вследствие катаболических эффектов гормонов. При сахарном диабете характерными являются истончение слизистой оболочки, сухость языка, парестезии, ангулярный хейлит [2].
Так, при гипофункции щитовидной железы отмечается сухость языка и губ, при ее гиперфункции — тремор, снижение вкуса [1, 11]. Утолщение языка и образование складок на нем (за счет увеличения количества нитевидных сосочков) в совокупности с другими симптомами могут иметь значение в ранней диагностике нарушения функции гипофиза [1]. При хронической недостаточности коры надпочечников на слизистой оболочке полости рта и боковых поверхностях языка появляются коричневые пятна различной формы без признаков воспаления. При гиперфункции коры надпочечников в полости рта могут быть выявлены афты, участки гиперкератоза, кандидомикоза вследствие катаболических эффектов гормонов. При сахарном диабете характерными являются истончение слизистой оболочки, сухость языка, парестезии, ангулярный хейлит [2].
Изменения в полости рта могут стать одними из ранних клинических симптомов заболеваний крови и органов кроветворения. Так, при анемии отмечается бледность слизистой оболочки, ее сухость, ангулярный стоматит, атрофия эпителия и сосочков языка, который становится острым и уменьшается по толщине [1, 7]. Особенностью гиперхромной анемии является избыточная увлажненность слизистой оболочки языка, появление гиперпигментированных пятен на небе, внутренней поверхности щек [2]. При гемофилии наблюдаются кровотечения из неповрежденных внешне десен и при прикусывании языка [1, 2]. При лейкозе можно обнаружить язвенно-некротические изменения слизистой оболочки полости рта [2].
Особенностью гиперхромной анемии является избыточная увлажненность слизистой оболочки языка, появление гиперпигментированных пятен на небе, внутренней поверхности щек [2]. При гемофилии наблюдаются кровотечения из неповрежденных внешне десен и при прикусывании языка [1, 2]. При лейкозе можно обнаружить язвенно-некротические изменения слизистой оболочки полости рта [2].
Поражения слизистой оболочки ротовой полости могут иметь место при инфекционных заболеваниях. В частности, при скарлатине характерным является малиновый язык (рис. 7).
У больных с сердечной недостаточностью могут появляться трофические изменения языка в виде инфильтрации, болезненности и некрозов. При декомпенсированных врожденных пороках сердца слизистая оболочка языка становится отечной, сухой, бледной с цианотичным оттенком [2, 11].
Для патологии почек характерными являются ксеростомия (сухость во рту), катаральный гингивит, эрозивный стоматит вследствие бактериального разложения мочевины слюной, так называемый аммиачный ожог [2, 11].
Одним из проявлений гиповитаминоза А является сухость слизистой полости рта, ороговение эпителия, при недостаточности витамина В1 — гиперплазия грибовидных сосочков, В6 — ангулярный стоматит, хейлит, при дефиците витамина В12 — десквамативный глоссит [2].
В качестве иллюстрации к вышеизложенному представляем следующий клинический случай. Ребенок Б., 7 лет, предъявлял жалобы на частый кашель в течение суток с трудноотделяемой мокротой. Из анамнеза заболевания известно, что кашель беспокоил мальчика в течение 4 месяцев, периодически с явлениями одышки, участием в акте дыхания вспомогательной мускулатуры, приступами свистящего дыхания с дистанционными хрипами. По результатам проведенного комплексного клинико-лабораторно-инструментального обследования был установлен диагноз: бронхиальная астма, атопическая форма, среднетяжелое течение, приступный период. В течение 1 мес. получал в качестве базисной терапии пульмикорт с незначительным эффектом и сохранением вышеуказанных жалоб. При оценке объективного статуса обращали на себя внимание выраженные фенотипические проявления дисплазии соединительной ткани: гипермобильность суставов, слабый мышечный тонус, сколиоз грудного отдела позвоночника, уплощенный свод стоп. Кожные покровы обычной окраски, чистые. Слизистая оболочка полости рта сухая, язык у корня обложен равномерным плотным желтым налетом, в области кончика языка справа очаг десквамации эпителия, искривление срединной линии в передней трети, деминерализованная зубная эмаль (рис. 8). Перкуторно над легкими ясный легочный звук, аускультативно в легких жесткое дыхание, хрипов нет. Другие органы и системы без патологии. При дополнительном сборе анамнеза выявлен положительный симптом «мокрой подушки», наличие частой отрыжки воздухом, что явилось основанием для исключения гастроэзофагеальной рефлюксной болезни, паразитарной инвазии.
Результаты проведенной фиброгастродуоденоскопии выявили умеренную гиперемию слизистой пищевода в нижней трети, пролабирование складок желудка до 2,5 см длины пищевода, слизистая оболочка желудка матовая, гиперемирована, с большим количеством слизи, примесью желчи. В фекалиях обнаружены цисты лямблий.
Учитывая результаты дополнительных методов исследования, к базисной терапии пульмикортом был добавлен мотилиум в течение 14 дней, назначено трехэтапное лечение лямблиоза. На фоне данного комплексного лечения отмечалась положительная динамика бронхообструктивного синдрома (уменьшение, а затем исчезновение кашля), состояния слизистой оболочки языка (отсутствие очага десквамации эпителия, повышение влажности слизистой оболочки, уменьшение налета и изменение его цвета) (рис. 9).
Таким образом, патологические изменения функций и структуры различных органов и систем проявляются изменениями слизистой оболочки языка начиная с ранних клинических стадий. Оценка состояния языка важна при описании объективного статуса пациента, особенно в динамике наблюдения. Ее отражение в медицинской документации является простым и важным скрининговым показателем для составления алгоритма диагностического поиска, а также дополнительным критерием эффективности лечения, реабилитации и профилактики различных заболеваний.
118. Развитие и строение языка. Сосочки языка, их строение и функциональное значение.
Развивается из корневых зачатков-миотомы образуют мышцу языка, а слизистая формируется за счет жаберных карманов. Срастаясь зачатки образуют язык. Язык это мышечное тело. Основу языка составляет поперечнополосатая мышечная ткань соматического типа. Снаружи язык покрыт слизистой оболочкой. Верхняя и боковая поверхности языка имеют неровный рельеф за счет сосочков языка и неподвижно сращены с мышечным телом. Подслизистая оболочка здесь отсутствует. Сосочки языка — выросты собственного слоя слизистой оболочки, покрытые снаружи многослойным плоским частично ороговевающим эпителием (нитевидные сосочки). Различают 4 вида сосочков: нитевидные, грибовидные, желобовидные, листовидные. Нитевидные сосочки — самые многочисленные, равномерно покрывают верхнюю поверхность языка, наиболее мелкие (длина 0,3 мм). При ряде заболеваний процесс отторжения поверхностных эпителиоцитов может замедляться, создавая картину языка, обложенного белым налетом. Грибовидные сосочки немногочисленные, располагаются на спинке языка и его кончике, края имеют форму гриба с узким основанием и широкой вершиной. Крупнее нитевидных В толще эпителия находятся вкусовые почки. Желобоватые сосочки (сосочки языка, окруженные валом) располагаются между телом и корнем языка. Отчетливо возвышаются над уровнем слизистой. Они имеют узкое основание и широкую, уплощенную свободную часть. Вокруг сосочка глубокая щель — желобок, отделяющий сосочек от валика — утолщения слизистой оболочки. В толще эпителия расположены вкусовые луковицы. Листовидные сосочки хорошо развиты у детей, располагаются по краям языка, в эпителии содержат вкусовые почки. В пространства, разделяющие листовидные сосочки, открываются выводные протоки слюнных белковых желез. Секрет желез промывает пространства между сосочками. Мышечная основа языка образует тело. Образована поперечнополосатой мышечной тканью, волокна которой идут в трех взаимно перпендикулярных направлениях: вертикально, продольно и поперечно. Слизистая оболочка на нижней поверхности покрыта многослойным плоским неороговевающим эпителием. Собственная пластинка вдается в эпителий, образуя короткие сосочки. Есть подслизистая основа.
Слюнные железы языка делятся на три вида: белковые, слизистые и смешанные. Белковые простые трубчатые разветвленные железы. Расположены поблизости от желобоватых и листовидных сосочков. Выводные протоки открываются в ровики сосочков, окруженные валиком, или между листовидными сосочками. Концевые отделы-разветвленные трубочки. Слизистые расположены в корне языка и вдоль его боковых краев. Одиночные простые альвеолярно-трубчатые разветвленные железы. Смешанные железы в переднем отделе. Их протоки открываются вдоль складок слизистой оболочки под языком. Секреторные отделы расположены в толще языка.
119. Строение зуба.
Основу каждого зуба образует дентин. Дентин зуба покрыт эмалью-это анатомическая коронка. Остальная часть покрыта цементом-это анатомическая шейка. Узкая часть между коронкой и корнем-шейка. Внутри зуба есть полость. Ее расширенная часть- пульпарная камера, заключающая ткань, богатую сосудами и нервами, называют пульпой зуба. Пульпарная камера переходит в узкий корневой канал. Зуб помещен в альвеолу, образованную костными гребешками, или альвеолярными отростками, нижней и верхней челюстей и прочно закреплен зубной или периодонтальной связкой. Коллагеновые волокна периодонта погружаются в костную альвеолу в виде шарпеевских волокон. Слизистая, покрывающая альвеолярные отростки, называется десной. ЧАСТЬ ЗУБА НАД ДЕСНОЙ-КЛИНИЧЕСКАЯ КОРОНКА.
Листовидные сосочки — это… Что такое Листовидные сосочки?
- Листовидные сосочки
вкусовые сосочки, расположенные по краям языка.
1. Малая медицинская энциклопедия. — М.: Медицинская энциклопедия. 1991—96 гг. 2. Первая медицинская помощь. — М.: Большая Российская Энциклопедия. 1994 г. 3. Энциклопедический словарь медицинских терминов. — М.: Советская энциклопедия. — 1982—1984 гг.
- Листериоз
- Листок нетрудоспособности
Смотреть что такое «Листовидные сосочки» в других словарях:
листовидные сосочки — (papillae foliatae, PNA, BNA, JNA) вкусовые сосочки, расположенные по краям языка … Большой медицинский словарь
Сосочки — Кластеры вкусовых почек, лежащие внутри небольших, но видимых возвышений на поверхности языка. Известны сосочки четырех типов, отличающиеся друг от друга формой и местоположением: грибовидные, нитевидные, листовидные и желобоватые … Психология ощущений: глоссарий
Полость рта — (cavum oris) (рис. 151, 156, 194) является началом пищеварительного аппарата. Спереди она ограничена губами, сверху твердым и мягким нёбом, снизу мышцами, образующими дно полости рта, и языком, а по бокам щеками. Открывается полость рта… … Атлас анатомии человека
Язык — I Язык (lingua) мышечный орган полости рта. В языке различают верхушку, тело и корень. У новорожденного язык короткий, широкий и толстый, целиком лежит в полости рта, его корень расположен горизонтально. По мере роста ребенка корень языка… … Медицинская энциклопедия
Язык — I Язык (lingua, или glossa) непарный вырост дна ротовой полости у позвоночных животных и человека. Я. рыб образован складкой слизистой оболочки; не имеет мускулатуры (за исключением двоякодышащих) и движется вместе со всем висцеральным… … Большая советская энциклопедия
Гортань — Проходя через носоглоточные отверстия, воздух поступает в верхнюю часть дыхательной трубки, которая называется гортанью (larynx) (рис. 156, 194, 202) и располагается в передней части шеи, под подъязычной костью, на уровне IV VII шейных позвонков … Атлас анатомии человека
Язык — Язык, lingua, мышечный орган, покрытый сверху, с боков и частично снизу слизистой оболочкой. В языке различают две части: переднюю, свободную, часть, или тело языка, corpus linguae, и заднюю корень языка, radix linguae. Тело языка, corpus… … Атлас анатомии человека
ЯЗЫК — (анат.), у наземных позвоночных и человека мышечный вырост (у рыб складка слизистой оболочки) на дне ротовой полости. Участвует в захвате, обработке пищи, в актах глотания и речи (у человека). На Я. расположены вкусовые рецепторы. Язык человека… … Естествознание. Энциклопедический словарь
Вкус — I (gustus) ощущение, возникающее при действии химических раздражителей (пищевых и непищевых веществ) на рецепторы языка и ротовой полости. С помощью В. эти вещества оцениваются по информационному (качественному и количественному) и гедоническому… … Медицинская энциклопедия
Язык (анатомия) — У этого термина существуют и другие значения, см. Язык (значения). Язык (анатомия) … Википедия
Пищеварительная система — обеспечивает усвоение организмом необходимых ему в качестве источника энергии, а также для обновления клеток и роста питательных веществ. Пищеварительный аппарат человека представлен пищеварительной трубкой, крупными железами пищеварительного… … Атлас анатомии человека
(PDF) Морфология и развитие листовых сосочков у Sematophyllaceae
Молекулярная систематика в UMSL. Мы также благодарим Роберта Мэджилла,
Брюса Аллена, Питера Стивенса, Билла Бак, Эндрю Дуст, Сару
Фуэнтес Сориано, Марка Бейльштейна, Элоизу Сари, Патрика Суини
и Жаклин Летт за их постоянную помощь и поддержку. Шэрон
Бартоломью-Беган и Анджела Ньютон также благодарит
за их ценные обзорные комментарии, которые значительно обогатили оригинальную версию
.
ЦИТИРОВАННАЯ ЛИТЕРАТУРА
Акияма, Х. и Н. Нисимура. 1993. Дальнейшие исследования ветки
почек у мхов; «Pseudoparaphyllia» и «чешуйчатые листья».
Journal of Plant Research 106: 101–108.
Boudier, P. 1990. Fissidens kozaninii Latzel en Touraine. E
´tude
de la papilosite
´du limbe. Cryptogamie Bryologie et
Liche
´nologie 11: 245–252.
Боззола, Дж. Дж. И Л. Д. Рассел.1998. Электронная микроскопия.
Принципы и методы для биологов. 2
-е издание
. Джонс
и Бартлетт, Садбери, Массачусетс.
Brotherus, V. F. 1925. Musci (Laubmoose). 2. Хальфте. В A.
Engler (ed.), Die natu
¨rlichen Pflanzenfamilien Ed.2.
Бак, В. Р. 1991. Основа семейной классификации
плеврокарпных мхов. Успехи в бриологии 4: 146–185.
———, К. Дж. Кокс, А. Дж. Шоу и Б.Goffinet. 2005. Порядковый номер
отношения плеврокарпных мхов, с особым акцентом на
Hookeriales. Систематика и биоразнообразие
2: 121–145.
——— и Б. Гоффине. 2000. Морфология и классификация
мхов. Страницы 71–123. В А. Дж. Шоу и Б. Гоффине,
Биология мохообразных. Издательство Кембриджского университета,
Кембридж.
——— & B. C. Tan. 1989 [1990]. Азиатские роды из
Sematophyllaceae, связанных с Trichosteleum.Acta
Bryolichenologica Asiatica 1: 5–19.
Ca
Маара, П. Е. А. С. 2006. Молекулярный вклад в систематическое размещение
мхов рода Paranapiacabaea.
Boletim do Instituto de Bota
ˆnica (Sa
˜o Paulo) 18: 159–162.
Кано, М. Дж. 1994. Importa
´ncia de la microscopı
´a eletro
´nica en
los estudios taxonomicos de brio
´fitos. Boletin de la
Sociedad Espan
˜ola de Briologı
´a 4: 1–3.
Casas De Puig, C. & L. Molinas. 1974. E
´tude au microscope
e
´letronique a
‘balayage de la surface des feuilles de Tortula
muralis (Hedw.) Gaertn. var. хирсута (Вентиляция) Пар. Ревю
Bryologique et Liche
´nologique 40: 267–270.
Кокс, К. Дж., Б. Гоффине, А. Дж. Шоу и С. Болес. 2004. Филогенетические
взаимоотношений между мхами на основе гетерогенного
байесовского анализа множественных генов из множественных геномных
сравнений.Систематическая ботаника 29: 234–250.
Де Луна, Э., У. Р. Бак, Х. Акияма, Т. Арикава, Т. Цубота,
Д. Гонза
´lez, А. Э. Ньютон и А. Дж. Шоу. 2000. Порядковый номер
Филогенияв пределах гипнобрилейных плеврокарпных мхов
выведен из кладистического анализа трех наборов данных последовательностей хлоропластной ДНК
: trnL-F, rps4 и rbcL. Бриолог
103: 242–256.
Гальего, М. Т. 2005. Таксономическое исследование рода Syntrichia.
Журнал ботанической лаборатории Хаттори 98: 47–122.
Гардинер, А., М. Игнатов, С. Хуттунен и А. Троицкий. 2005. На
воскресение семейств Pseudoleskeacae Schimp. и
Pylaisiaceae Schimp. (Musci, Hypnales). Таксон 54:
651–663.
Гоффине Б., К. Дж. Кокс, А. Дж. Шоу и Т. А. Дж. Хеддерсон. 2001.
Bryophyta (мхи): систематические и эволюционные
выводы из филогении гена rps4 (хпДНК).Анналы
Ботаника 87: 191–208.
Герра, Дж., Р. М. Рос и Дж. С. Каррио
´n. 1992. Таксономический статус
Tortula muralis var. baetica (Musci, Pottiaceae) — сравнительное исследование
. Журнал бриологии 17: 275–283.
Hedena
¨s, L., B. C. Tan & C. Ying. 2007. [Abstract]
Морфологические и молекулярные
предполагают различные отношения между видами Acroporium Mitt. и близко
родственных родов (Sematophyllaceae).Стр. 30. В Анналах
Всемирная конференция бриологов. Куала Лумпур.
Huttunen, S., M. S. Ignatov, K. Mu
¨ller & D. Quandt. 2004.
Филогения и эволюция эпифитизма у трех семейств мхов
: Meteoriaceae, Brachytheciaceae и
Lembophyllaceae. Монографии по систематической ботанике из
Ботанический сад Миссури 98: 328–361.
Игнатов М.С. и Хедена Л.
¨s. 2007. Гомологии стволовых
структур плеврокарпных мхов, особенно
псевдопарафиллий и подобных структур.В A. E. Newton
и R. S. Tangney (ред.), Pleurocarpous mossessystematics
и эволюция. Специальный том 71 ассоциации систематики:
269–286.
Ирландия, Р. Р. 1991. Кутикулярные сосочки Atrichum crispum. Бриолог
94: 73–76.
La Farge-England, C. 1996. Форма роста, схема ветвления,
и перихециальное положение во мхах: кладокарпия и
плеврокарпия пересмотрены. Бриолог 99: 170–186.
Мэджилл, Р.E. 1977 г. Повторное исследование Globulinella (Musci;
Pottiaceae). Бриолог 80: 76–82.
———. 1990. Glossarium polyglottum bryologiae: многоязычный глоссарий
по бриологии. Монографии в систематической ботанике
из ботанического сада Миссури 33: 1–297.
———, Ф. Сибери и Д. М. Дж. Мюллер. 1974. Оценка метода сушки критической точки
и его применения в систематических исследованиях мхов
. Бриолог 77: 628–632.
Мишлер Б. Д. 1986. Онтогенез и филогения в Тортуле
(Musci: Pottiaceae). Систематическая ботаника 11: 189–208.
———. 1987. Развитие листьев у Tortula papillosissima
(Pottiaceae). Воспоминания о ботаническом саду Нью-Йорка
45: 48–54.
———. 1988. Взаимосвязь онтогенеза и филогении,
применительно к мохообразным. Страницы 117–136. В C. J.
Хамфрис (ред.), Онтогенез и систематика. Колумбия
University Press, Нью-Йорк.
Мозинго, Х. Н. П., Ю. Кляйн, Ю. Зееви и Э. Р. Льюис. 1969.
Исследования Sphagnum
inbricatum с помощью сканирующего электронного микроскопа. Бриолог 72: 484–488.
Ca
ˆmara & Kellogg: Сосочки листьев у Sematophyllaceae 31
В Halophyte Seashore Paspalum для секвестрации натрия используются адаксиальные папиллы листьев
Abstract
Во всем мире прогнозируется растущая проблема засоленности, при этом почти 30% земель будут потеряны из-за засоления почвы в ближайшие 30 лет.Многие злаковые культуры, жизненно важные для поддержания мирового потребления калорий, чувствительны к соли. Изучение механизмов солеустойчивости галофитных трав, растений, которые процветают в солевых условиях, может быть эффективным подходом для улучшения в конечном итоге солеочувствительных травяных культур. Seashore paspalum ( Paspalum vaginatum ) — галофитная трава Panicoid, способная расти в концентрациях солей, близких к концентрации солей в морской воде. Несмотря на его широко распространенное культивирование в качестве устойчивого газонного травяного покрова, механизм, лежащий в основе его способности сохранять высокие концентрации Na + в фотосинтетической ткани при сохранении роста, остается неизвестным.Мы исследовали структуру листьев и содержание ионов у P. vaginatum ‘HI10’, который демонстрирует повышенный рост в условиях солевого раствора, и Paspalum distichum ‘Spence’, который демонстрирует замедленный рост в условиях соли, чтобы лучше понять превосходную солевую толерантность cv HI10. Поразительным различием между cv HI10 и cv Спенс был высокий стабильный уровень K + в cv HI10. Визуализация также показала, что адаксиальная поверхность как cv HI10, так и cv Spence содержала плотные реберные гребни сосочков.Однако эти одноклеточные отростки эпидермиса были значительно больше у сорта HI10, чем у сорта Спенс. Было показано, что сосочки cv HI10 действуют как поглотители Na + при выращивании растений в засоленных условиях. Мы предоставляем доказательства того, что сосочки листа функционируют как специализированные структуры для секвестрации Na + в P. vaginatum , иллюстрируя возможный путь биотехнологического улучшения солеочувствительных культур Panicoid с аналогичными структурами листьев.
Около 20% орошаемых земель считаются засоленными, причем количество засоленных почв во всем мире увеличивается (Mayak et al., 2004). Это связано с увеличением орошения сельскохозяйственных полей, вызванным более частыми засухами из-за изменения климата. Эта тенденция вызывает тревогу из-за высокой солевой чувствительности большинства видов сельскохозяйственных культур, которые являются жизненно важными для нас ресурсами. Снижение урожайности сельскохозяйственных культур на засоленных почвах приводит к потерям от 12 до 27,3 миллиардов долларов США ежегодно (Qadir et al., 2014). Таким образом, улучшение солеустойчивости растений станет ключевым фактором в ближайшие десятилетия. Выращивание солеустойчивых культур — это рентабельный подход к улучшению роста на засоленных почвах.Хотя большая часть работы была сосредоточена на разведении солеустойчивых видов, прогресс в этой области был медленным из-за сложной генетической и физиологической природы солевой реакции. Более того, большинство исследований проводилось на гликофитных модельных системах, чувствительных к соли (Munns and Gilliham, 2015). Поэтому выявление механизмов солеустойчивости у галофитов, видов, которые могут завершить свой жизненный цикл при концентрациях соли 200 мм, и передача этих путей в гликофиты представляет большой интерес (Rajalakshmi and Parida, 2012; Roy and Chakraborty, 2014).
И гликофиты, и галофиты развили множество механизмов солеустойчивости, включая исключение, секвестрацию и секрецию натрия (Na + ); производство осмолита; ионный гомеостаз; и детоксикация активных форм кислорода (АФК) (Meng et al., 2018). Часто механизмы, присутствующие в гликофитах, такие как продукция осмолита и исключение Na + , используются в галофитах с более высокой эффективностью (Wyn Jones and Storey, 1981; Grieve and Maas, 1984). Однако галофиты также используют механизмы, отсутствующие у гликофитов.Секвестрация и секреция соли через солевые железы — это специфический для галофитов механизм борьбы с солью (Flowers and Colmer, 2008). Солевые железы обнаружены у более чем 50 видов в 14 семействах покрытосеменных с четырьмя подтипами: клетки эпидермального пузыря, сложные многоклеточные железы, двухклеточные железы и одноклеточные железы (Dassanayake and Larkin, 2017). Отряд Поалеса содержит ~ 8% всех галофитов (Flowers et al., 2010) и, следовательно, был в центре внимания многих работ, посвященных солевым железам (Ceccoli et al., 2015).Поскольку солеустойчивость независимо эволюционировала более чем в 70 раз у линий трав (Bennett et al., 2013), изучение этих секвестрирующих / секретирующих солевых структур в травах является отличным подходом для лучшего понимания механизмов солеустойчивости у галофитов.
Большая часть структурных и физиологических исследований солевых желез трав была проведена в подсемействах Chloridoideae и Oryzoideae. Травы несут на поверхности листьев одноклеточные или двухклеточные железы, часто называемые железистыми трихомами или микроволосами (Dassanayake and Larkin, 2017).Микроволосы наблюдались на поверхности листьев у всех подсемейств трав, кроме Pooideae, и в процессе эволюции они приобрели разнообразные функции, включая секвестрацию или секрецию таких веществ, как каллоза и тяжелые металлы (Burke et al., 2000; Ceccoli et al., 2015). Одноклеточные структуры на адаксиальной стороне листа, способные секретировать соль, обнаруживаются только у диких видов риса Oryzoideae Porteresia coarctata (Flowers et al., 1990; Sengupta and Majumder, 2009). Солевые железы у Chloridoideae двухклеточные, состоящие из крышечной клетки и нижней базальной клетки, обе из которых плотны в цитоплазме и митохондриях (Ceccoli et al., 2015). У некоторых видов кутикула утолщена над крышкой клетки, образуя кутикулярную камеру, используемую для хранения секретируемых солей (Amarasinghe and Watson, 1988). У Panicoideae сообщалось о нескольких случаях секреции Na + (McWhorter et al., 1995; Ramadan and Flowers, 2004), но на сегодняшний день никаких структур секвестрации выявлено не было.
Подсемейство Panicoideae включает в себя важные с агрономической точки зрения продовольственные культуры кукурузу ( Zea mays ) и сорго ( Sorghum bicolor ) в дополнение к биотопливным травам мискантус ( Miscanthus sinensis ), просо прутьевому ( сахар Panicum ). ( Saccharum officinarum ).Одним из наиболее солеустойчивых видов Panicoideae является галофит морского побережья paspalum ( Paspalum vaginatum ). Его выращивают во всем мире как газон, и его популярность объясняется его способностью орошаться солоноватой водой. P. vaginatum может выжить при концентрациях соли, близких к концентрации морской воды (Uddin et al., 2012), и использует продукцию осмолита, ионный гомеостаз и исключение Na + , чтобы справиться с солевым стрессом (Peacock and Dudeck, 1985; Lee et al., др., 2008; Guo et al., 2016). Однако его способность поддерживать рост при накоплении высоких уровней Na + в тканях листа вызывает недоумение.
Здесь мы изучили структуру листьев и секвестрацию Na + у сорта «HI10», сорта P. vaginatum и сорта «Spence», сорта Paspalum distichum . P. vaginatum и P. distichum являются близкородственными (и, возможно, одним и тем же видом; Eudy et al., 2017) и составляют группу «Disticha» в трибе Paspaleae. P. distichum менее солеустойчив, чем P. vaginatum , и обычно встречается в пресноводных местообитаниях (Leithead et al., 1971). Таким образом, P. vaginatum и P. distichum представляют собой полезную пару видов для изучения солеустойчивости. Кроме того, их солевые реакции можно сравнить с реакциями сорго, паникоидного гликофита. Основная цель нашего исследования заключалась в определении фенотипических и физиологических факторов, которые способствуют дифференцированной устойчивости к солевому стрессу двух видов Paspalum spp.сорта и сорго «BTx623». Мы показываем, что оба вида Paspalum содержат плотные ряды полупрозрачных сосочков на адаксиальной поверхности. Сосочки представляют собой одноклеточные выступы из клеток эпидермиса и намного больше у сорта HI10, чем у сорта Спенс. Мы также демонстрируем, что сосочки секвестируют Na + при солевом стрессе. Таким образом, это исследование предоставляет доказательства секвестрации Na + в специализированных листовых органах внутри Panicoideae.
РЕЗУЛЬТАТЫ
cv HI10 и cv Spence различаются по солевой реакции при 0, 10 и 30 dS м
−1 Уровни солиP.vaginatum «HI10» усиливает рост в соленой воде по сравнению с пресной водой, в то время как у сорта Спенс рост замедляется. Поведение роста сорта Спенс при солевом стрессе аналогично тому, что обычно наблюдается у гликофитов, таких как сорго (). cv HI10 и cv Spence также демонстрируют различную морфологию листьев, при этом cv Spence имеет более крупные листья с грубой текстурой по сравнению с cv HI10 ().
Реакция роста растений на различные уровни солевого стресса. От A до C: реакция роста была измерена для S.bicolor ‘BTx623’ (A), P. distichum ‘Spence’ (B) и P. vaginatum ‘HI10’ (C). Уровни соли даны как проводимость в децисименсах на метр. Репрезентативные растения для каждого уровня соли фотографировали через 6 недель роста при 0 (пресная вода), 10 или 30 dS м -1 соли. D, Морфология листьев сортов Спенс (вверху) и HI10 (внизу). Масштабная линейка = 1,3 см.
Сравнение уровней Na + в над- и подземных органах cv HI10, cv Spence и cv BTx623 выявило значительные различия в количестве, а также в распределении Na + между двумя видами Paspalum и cv BTx623. (; Дополнительный рис.S1, A – C). Во всех органах средние уровни Na + ниже 30 децисименс (dS) м -1 солевого стресса составляли 44 968 ppm для cv BTx623, 14 917 ppm для cv HI10 и 12 175 ppm для cv Спенс. При солевом стрессе 30 dS м -1 cv BTx623 накапливал наибольшее количество Na + в ткани ствола, в то время как самые высокие уровни Na + у cv HI10 и cv Spence наблюдались в корнях (). Внутри Paspalum spp. ткань листа, cv HI10 секвестрировала значительно большее количество Na + в оболочке листа по сравнению с листовой пластинкой под 10 и 30 dS м -1 соли ().Та же тенденция наблюдалась и у сорта Спенс, хотя разница не была статистически значимой.
Уровни ионов в различных органах видов Paspalum и S. bicolor . Содержание Na + (A – C), содержание K + (D – F) и соотношение K + / Na + (G – I) определяли в различных фракциях органов у сорго сорта BTx623 (A , D и G), P. distichum ‘Spence’ (B, E и H) и P. vaginatum ‘HI10’ (C, F и I) после 6 недель роста при разных уровнях соли. .Значения представлены как среднее ± стандартное отклонение для трех образцов на образец на обработку. Буквы в нижнем регистре над каждой полосой указывают на статистическую значимость на уровне P <0,05, на что указывает односторонний анализ ANOVA с тестом разделения средних значений по методу достоверно значимой разницы (HSD) Тьюки, проводимому в пределах каждого генотипа по типу иона.
Содержание калия (K + ) (; дополнительный рис. S1, D – F), которое в среднем составляло 22 780 ppm по органам у cv BTx623, 34 767 ppm у cv HI10 и 19 428 ppm у cv Спенс под пресной водой, значительно увеличилось в надземные органы в cv BTx623 при 30 dS м -1 соли (), но оставались в основном постоянными для cv HI10 во всех органах, кроме столонов (), где значительное снижение содержания K + наблюдалось под солью.У сорта Спенс концентрация K + была значительно ниже во всех надземных тканях у растений, выращенных под 30 dS м -1 соли по сравнению с пресной водой ().
Во всех органах соотношения K + / Na + в пресной воде были сопоставимы для cv BTx623 и cv Spence, со средними отношениями 4625,1 и 2419,7 соответственно, но были на два порядка ниже для cv HI10 (среднее значение 61,8;). Соотношения были постоянно выше в надземных тканях по сравнению с корнями (дополнительный рис.S1, G – I). Солевой стресс привел к значительному снижению отношений K + / Na + у всех образцов, но кратность изменения была намного выше у сортов BTx623 и Спенс, чем у сорта HI10. Фактически, хотя под пресной водой отношения K + / Na + значительно ниже, в надземных тканях растений, выращенных под 30 dS m -1 соли, были выше у сорта HI10 (в среднем 3,1), чем у любого сорта BTx623 ( в среднем 1,0) или сорта Спенс (в среднем 1,8; дополнительный рис. S1, G – I).
cv HI10 и cv Spence демонстрируют отчетливую морфологию поверхности листа
Для дальнейшего изучения различных закономерностей накопления соли у cv HI10, cv Spence и cv BTx623 при солевом стрессе мы сравнили анатомию их листьев. Световая микроскопия показала, что структура листового влагалища была сходной у сортов HI10, сортов Спенс и сортов BTx623 (). Однако в пластинке листа одноклеточные протяженные эпидермальные клетки, называемые сосочками, наблюдались над каждой крупной продольной жилкой у Paspalum spp.(), но не в cv BTx623 ().
Поперечные изображения влагалища листа и листовой пластинки. Поперечные срезы влагалища (A, C и E) и листовой пластинки (B, D и F) у P. vaginatum ‘HI10’ (A и B), P. distichum ‘Spence’ (C и D), и сорго cv BTx623 (E и F), выращенное в пресной воде, получали изображения под световым микроскопом. Масштабные линейки = 50 мкм.
Диаметр абаксиальных и адаксиальных эпидермальных клеток достоверно не различается между обработкой солью у Paspalum spp.генотипов ни в cv BTx623 (). Средний диаметр абаксиальных клеток был значительно выше у cv HI10 по сравнению с cv Spence и cv BTx623, составляя в среднем 23,0 мкм для разных обработок по сравнению с 19,2 и 20 мкм для cv Spence и cv BTx623, соответственно (). Значительные различия в диаметре адаксиальных клеток также наблюдались между сортами HI10, сорта Спенс и сортами BTx623, при этом CV HI10 составлял в среднем 32,3 мкм при разных обработках по сравнению с 24,8 мкм и 21,5 мкм у сорта Спенс и сорта BTx623, соответственно (). Различия в размере эпидермальных клеток на адаксиальной стороне листа были еще более поразительными, когда только клетки сосочка (шесть верхних клеток над каждой основной жилкой) были измерены в cv HI10 и cv Spence, при среднем диаметре клеток 37.1 мкм у сосочка cv HI10 по сравнению с 20,1 мкм у сорта Спенс при разных обработках (). Количество эпидермальных клеток над каждой основной веной было одинаковым для cv HI10 и cv Spence, в среднем 8,3 клеток и 7,6 клеток на основную вену при лечении cv HI10 и cv Спенс, соответственно ().
Диаметр абаксиальных и адаксиальных клеток эпидермиса. На графиках показаны диаметры абаксиальных эпидермальных клеток (A), адаксиальных эпидермальных клеток (B), клеток сосочка (C) и количество адаксиальных клеток над каждой основной жилкой (исключая устьичные и булевидные клетки; D) в листьях сорго cv BTx623 , с.distichum ‘Spence’ и P. vaginatum ‘HI10’, выращенные в течение 6 недель под 0 (пресная вода), 10 и 30 dS м -1 соли. Диаметр клеток сосочков определяли путем измерения шести самых верхних клеток над каждой основной веной. Значения представлены в виде среднего ± стандартная ошибка для n = 162–227 клеток (диаметр абаксиальных клеток), 199–260 клеток (диаметр адаксиальных клеток), 72–100 клеток (диаметр сосочковых клеток) и 18–24 основных клеток. вены, используемые для подсчета количества клеток. Строчные буквы над каждой полосой указывают на статистическую значимость на уровне P <0.05, на что указывает однофакторный анализ ANOVA с тестом разделения средних значений по методу достоверной значимой разницы Тьюки (HSD) по типам клеток для разных образцов и обработок солью.
Изображения, полученные с помощью мощного препаровального микроскопа, показали, что сосочки полупрозрачные и располагаются рядами по всей адаксиальной поверхности листа (). Сканирующая электронная микроскопия (sem) подтвердила, что сосочки были значительно больше у сорта HI10, чем у сорта Spence (). СЭМ-изображения двух дополнительных P.vaginatum , cv 509018-3 и cv KC9, показали, что у обоих были большие сосочки, которые были подобны по размеру таковым у cv HI10 (дополнительный рис. S2, A – D). Изображения SEM также показали, что реберные гребни с сосочками перекрывают бороздки, содержащие устьицы (дополнительный рис. S2E).
Сосочки на адаксиальной стороне листа влияют на гидрофобность листа. От A до D, изображения адаксиальной поверхности листа P. с помощью диссекционного микроскопа (A и B) и сканирующего электронного микроскопа (C и D).distichum «Spence» (A и C) и P. vaginatum «HI10» (B и D), выращенные в пресной воде. E. Репрезентативные изображения статических капель воды на адаксиальной и абаксиальной поверхностях листа показаны для cv HI10, cv Spence и cv BTx623. Масштабные линейки = 100 мкм. Рабочее расстояние SEM составляло 10,3 мм для cv Spence (C) и 10,4 мм для cv HI10 (D) с увеличением в 1000 раз для обоих.
Абаксиальные поверхности cv HI10, cv 509018-3, cv KC9 и cv Spence несут микроволокнистые структуры (дополнительный рис.S3, A – D), которые морфологически сходны с двухклеточными трихомами, обнаруженными у паникоидной травы Johnsongrass ( Sorghum halepense ; McWhorter et al., 1995). Их плотность была значительно выше на абаксиальной поверхности у сорта Спенс, чем у трех образцов P. vaginatum (дополнительный рисунок S3F). Микроволосы также наблюдались на адаксиальной поверхности у сорта Спенс (дополнительный рис. S3E). Они не были замечены на адаксиальной поверхности у трех представленных образцов P. vaginatum , хотя возможно, что они присутствовали, но были скрыты большими сосочками.Плотность адаксиальных микроволокон у сорта Спенс была аналогична плотности абаксиальных микроволокон у трех образцов P. vaginatum . Никаких существенных различий в плотности между пресноводными и солеными обработками не наблюдалось (дополнительный рис. S3F).
У сорта Спенс плотность устьиц была значительно выше на адаксиальной стороне, чем на абаксиальной (дополнительный рис. S3G). Хотя адаксиальные устьицы также присутствовали у образцов P. vaginatum , как можно увидеть на поперечных срезах листа (), они в основном были скрыты большими сосочками на изображениях SEM и, следовательно, не учитывались.За исключением сорта 509018-3, у которого наблюдалось увеличение абаксиальной плотности устьиц при солевом стрессе, не наблюдалось значительных различий в плотности устьиц между обработкой пресной водой и соленой водой (дополнительный рисунок S3G).
Измерения угла смачивания: прокси для размера сосочка
Чтобы определить, влияет ли размер сосочка на гидрофобность поверхности листа, мы измерили угол смачивания капель воды, помещенных на адаксиальную и абаксиальную поверхности листа cv HI10, cv Spence и cv BTx623.Адаксиальные поверхности листьев, содержащие сосочки, были значительно более гидрофобными, чем абаксиальные поверхности у сортов HI10 и Спенс (;). У cv BTx623, у которого отсутствуют сосочки, обе поверхности показали одинаковый уровень гидрофобности (;). Адаксиальная поверхностная гидрофобность была значительно выше у cv HI10, чем у cv Spence, и оба имели значительно более высокую адаксиальную гидрофобность, чем cv BTx623. Напротив, не было различий в гидрофобности абаксиальной поверхности листа между сортами HI10 и Спенс, а только незначительные различия между сортами BTx623 и двумя образцами Paspalum .Измерения адаксиальной и абаксиальной гидрофобности существенно не различались между листьями из Paspalum spp. растения, выращенные под пресной водой (0 dS m -1 ) и 30 dS m -1 соли (дополнительная таблица S1).
Таблица 1.
Измерения среднего краевого угла смачивания статических капель воды на адаксиальной и абаксиальной поверхностях листаПоказанные значения представляют собой средние значения ± se. Строчные буквы обозначают статистическую значимость на уровне P <0.05, на что указывает однофакторный анализ ANOVA с использованием критерия разделения среднего значения по методу достоверно значимой разницы Тьюки (HSD). n = 9–15 измерений на образец на обработку.
| Сорт (виды) | Адаксиальный контактный угол | Абаксиальный контактный угол | |||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ‘HI10’ ( P. vaginatum ) | 124.6532 33 | 124.6532 ± 1.77 a | ‘PI 299042’ ( P. vaginatum) | 110.08 ± 2,60 b | 63,55 ± 1,32 d | ||||||||||||||||||||||||||||||||||||||||||||
| ‘Tropic Shore’ ( P. distichum) | 114,38 ± 0,86 b | 64,12 ± 1,23 d | |||||||||||||||||||||||||||||||||||||||||||||||
| ‘Spence distichum () | 101,76 ± 1,20 c | 65,20 ± 1,39 d | |||||||||||||||||||||||||||||||||||||||||||||||
| ‘BTx623’ ( S. bicolor ) | 61,80 ± 1,18 d | 60,81 ± 1,30 d | 905 коррелированный с размером сосочка измерения краевого угла смачивания проводили на двух дополнительных образцах, P.vaginatum «PI 299042» и P. distichum «Tropic Shore» (). Кроме того, мы измерили ширину минимум 100 сосочков у этих образцов, а также у cv HI10 и cv Spence, по изображениям, полученным при препарировании под микроскопом (дополнительный рис. S4). cv PI 299042 и cv Tropic Shore имеют размер и текстуру листьев, аналогичные таковым у сорта Spence, а cv PI 299042 действует так же, как cv Spence при солевом стрессе (дополнительный рисунок S5). Хотя cv PI 299042 считается принадлежащим к P.vaginatum , а сорта Spence и Tropic Shore были классифицированы как P. distichum , анализ генотипа с использованием маркеров простого повтора последовательности (SSR) показал, что три образца принадлежали к одной и той же генетической субпопуляции в группе Disticha. cv HI10 принадлежал к другой генетической группе населения (Eudy et al., 2017). Сосочки у сорта PI 299042 и сорта Tropic Shore были сопоставимы по размеру, но были больше, чем у сорта Спенс, и меньше, чем у сорта HI10 (дополнительный рис.S4A). Адаксиальная поверхностная гидрофобность у четырех Paspalum spp. Образцы были достоверно положительно коррелированы со средней шириной ячеек сосочка ( R = 0,98, P = 0,0162).
ВКУСОВЫЕ НАБОРЫ И КЛЕТКИ
Вкусовые рецепторы находятся в основном в сосочках языка.Язык содержит 4 типа сосочков, самый распространенный тип, нитевидные, — тонкие и проволочные. shape и не содержат вкусовых рецепторов. На дорсальной, передней границе на языке грибовидные сосочки, грибовидные, имеют вкус бутоны расположены недалеко от середины или в расщелине сосочков. листовидных сосочков имеют форму листа со вкусовыми сосочками сбоку от сосочков, и это по границе. округлых сосочков содержат вкусовые рецепторы вдоль стороны мутовок и расположены в задней трети языка у форма V. Вкусовые рецепторы также расположены в слизистой оболочке ротовой полости нёбо и надгортанник.
Вкусовые клетки представляют собой модифицированные эпителиальные клетки, которые функционируют как сенсорные рецепторы.Около 50-60 вкусовых клеток расположены во вкусовых сосочках грушевидной формы. и вкусовые клетки через микроворсинки выступают во вкусовые поры. Там нерецепторные базальные клетки, которые расположены на базальной мембране которые не выступают во вкусовые поры. Эти базальные клетки дифференцируются через серию морфологических шагов в зрелую вкусовую клетку. В вкусовые клетки заменяются примерно каждые 10 дней. | |||||
Рисунок 8 | |||||
| Раньше считалось, что вкусовые рецепторы для сладкого существовали только на кончике языка (рис.9) и для горького (рис. 10) находились в заднем отделе, но в настоящее время известно что язык вообще не имеет региональных отличий на вкус качества. Кроме того, вкусовые клетки могут реагировать на раздражители самых разных молекулы вкуса сладкие, соленые, кислые или горькие. |
|
СЕНСОРНАЯ ПЕРЕДАЧА ВКУСА
Преобразование химических раздражителей в нервные импульсы начинается со слюны транспортировка растворенных полярных молекул, которые являются сладкими или горькими i.е. сахар, кофеин по вкусу (рис.12). Полярные молекулы do не проникают во вкусовые клетки, но связываются с рецепторами микроворсинок в вкусовые поры. Эти рецепторы связаны с G-белком, который называется густдуцин . с последующей активацией мессенджера 2 nd , который управляет лиганд-зависимый белок канала для уменьшения оттока K + через белок ионно-управляемого канала в клеточной мембране.Рецепторный потенциал генерируется, который открывает зависимый от напряжения канал Ca ++, и последующий разряд нейромедиатора в синапс. Затем в окончании афферентного нерва генерируется потенциал действия . и проводится стимулированным черепным нервом в ЦНС. Электролиты , такие как соли или кислоты, непосредственно взаимодействуют с рецептором вкуса клеточная мембрана, чтобы открыть белок ионно-закрытого канала и увеличить поток Na + и K +. Неполярные или липофильные стимулы переносятся к вкусовой клетке, но связывают с белком, связывающим белок, возможно продуцируются небольшими серозными железами, железами фон Эбнерса. Аналогичная трансдукция процесс преобразует неполярные растворенные вещества в нервные импульсы. Многие наркотики неполярные растворенные вещества, а некоторые имеют горький вкус. | |
Рисунок 11 |
Рисунок 12 |
ТРАНСМИССИЯ И ВОСПРИЯТИЕ ВОЗМОЖНОСТИ ДЕЙСТВИЯ КАК НЕЙРОННЫЙ КОД
Нейронные сигналы от одного вкусового рецептора не передаются напрямую через один аксон в центральную нервную систему вместо множества вкусовых клеток во многих вкусовых рецепторах стимулируется разнообразным качеством пережеванной пищи.Одна ветвь лицевого нерва широко разветвляется. и расходятся на несколько сосочков и на разные вкусовые рецепторы в пределах одного сосочки. Стимуляция сотен вкусовых рецепторов на языке порождает нейронный код , специфичный для каждого типа молекул растворенного вещества, которые может иметь сладкий, горький, соленый или кислый вкус. Люди могут обнаруживать, различать и определить многие вещества, обладающие особыми вкусовыми качествами, потому что нейронный код для каждого типа вкусовых стимулов декодируется центральной нервной системой. система.
| Черепные афферентные нервные волокна, VII, IX, X, выступают из язык в солитарное ядро в синапс с 2 nd порядок сенсорных нейронов. Волокна из ядра solitarius восходят к таламус и расходятся к парабрахиальному ядру. От таламуса, нерва волокна выходят на вкусовую кору и от парабрахиального ядра, волокна передаются в гипоталамус и миндалину. | |
Рисунок 13 |
НАРУШЕНИЯ ВКУСА
Дефицит вкусовых ощущений может быть вызван возрастом, наркотиками, болезнями, или травма. Например, с возрастом людей количество сосочков, содержащих вкусовые рецепторы, и снижение чувствительности ко многим вкусовым ощущениям растворенные вещества; однако пожилые люди по-прежнему любят сладкие, вкусные блюда.
Роль слюны во вкусовом восприятии очень важна, потому что слюна помогает в растворении растворенных веществ и в транспортировке их к поре вкуса. Это трудно что-либо попробовать, если во рту сухо. Несколько препаратов уменьшают слюноотделение течет и заставит стоматологов жаловаться на сухость во рту. Пациенты, которые получить лучевую терапию при раке полости рта, если у вас серьезный дефицит вкусовых качеств потому что лечение вызывает атрофию слюнных желез и снижает количество сосочков языка.Эти пациенты будут жаловаться на болезненные ощущения во рту и человек. потеря вкуса.
Причины нарушения вкуса
|
О дефиците вкуса сообщают пациенты с дефицитом витамина А, диабетом, гипотиреоз, почечная недостаточность и аутоиммунные заболевания, такие как Sjogren синдром.Нервная травма одного из афферентных черепных нервов из-за вкуса, например лицевого нерва, нарушит вкусовые ощущения и аромат продуктов. Распространенной острой потерей вкуса является контакт горячей жидкости с тканями полости рта. который повреждает сосочки и вкусовые рецепторы, к счастью, восстановление поврежденных вкусовые рецепторы обычно возникают в течение 10 дней.
Контент предоставлен Dr.Макс Хатчинс
Авторские права © 2001, Все права защищены
Научный центр здравоохранения Техасского университета в Хьюстоне (UTHealth)
Создано сотрудниками Мультимедиа
Скрипторий
Plantae | Галофит приморский паспалум использует адаксиальные сосочки листьев для связывания натрия (Plant Physiol.)
Глобальное потепление и интенсивное земледелие увеличивают засоление почвы, которое, по прогнозам, затронет 30% обрабатываемых земель к 2050 году. Засоление представляет собой серьезную угрозу для сельского хозяйства, как и большинство других посевы резко снижают свой рост и продуктивность в засоленных средах.Понимание стратегий, используемых галофитами (солеустойчивыми растениями), и их передача гликофитам (солеустойчивым растениям) может быть средством повышения урожайности сельскохозяйственных культур на засоленных почвах. Например, раскрытие механизмов, лежащих в основе способности солеустойчивого Paspalum vaginatum выживать вблизи морского побережья, может быть полезным для улучшения продуктивности солеочувствительной кукурузы и сорго, принадлежащих к одному подсемейству (Panicoideae), в ответ на высокая концентрация соли.Спикерман и Девос исследовали дифференциальную реакцию на засоление двух близкородственных видов паникоидов — P. vaginatum ‘HI10’ и P. distichum ‘Spence’, демонстрирующих повышенный и пониженный рост в условиях солевого раствора соответственно. Оба образца содержат плотные ряды сосочков на адаксиальном эпидермисе листа. Однако подробный морфологический анализ показал, что «HI10» образует более крупные сосочки. Помимо предполагаемой функции опускания Na + , эти большие эпидермальные структуры могут также регулировать потерю воды через транспирацию, скрывая устьица.Авторы использовали флуоресцентный краситель CoroNa Green для измерения содержания Na + в очищенных сосочках и обнаружили значительное увеличение флуоресценции у HI10, но не у Spence, в ответ на солевую обработку, что указывает на связь между солеустойчивостью. и способность связывать Na + в специализированных эпидермальных структурах листа. Это исследование открывает путь к биотехнологическому применению механизмов, специфичных для галофитов, для улучшения солеустойчивости сельскохозяйственных культур. (Резюме и адаптация изображения Микелы Оснато @michela_osnato) Физиология растений.10.1104 / стр. 20.00796
| [1] | Вэньбинь Чжоу, Цю-Юнь (Дженни) Сян и Цзюнь Вэнь. Филогеномика, биогеография и эволюция морфологии и экологической ниши восточной Азии — востока Северной Америки Нисса (Nyssaceae) [J]. J Syst Evol, 2020, 58 (5): 571-603. |
| [2] | Виктор Люсия, М.Монтсеррат Мартинес-Ортега, Энрике Рико и Кесара Анамтават-Йонссон. Открытие рода Pseudoroegneria (Triticeae, Poaceae) в Западном Средиземноморье при изучении родовых границ Elymus [J]. J Syst Evol, 2019, 57 (1): 23-41. |
| [3] | Стефани М. Икерт-Бонд, Эй Джей Харрис, Сью Лутц, Джун Вен.Детальное изучение микроморфологии и анатомии листьев Нового Света Vitis L. подрод Vitis в рамках филогенетических и экологических рамок показывает эволюционную конвергенцию [J]. J Syst Evol, 2018, 56 (4): 309-330. |
| [4] | С. Робберт Градштейн, Руи-Лян Чжу, Лей Шу, Альваро Х. Перес. Reinerantha foliicola , новый род и вид Lejeuneaceae subtribe Cololejeuneinae (Marchantiophyta) из Эквадора [J].J Syst Evol, 2018, 56 (1): 67-75. |
| [5] | Роберт Дж. Сорен, Пол М. Петерсон, Константин Ромащенко, Геррит Давидсе, Джордан К. Тейшер, Линн Дж. Кларк, Патрисия Барбера, Линн Дж. Гиллеспи, Фернандо О. Сулоага. Всемирная филогенетическая классификация Poaceae (Gramineae) II: обновление и сравнение двух классификаций 2015 г. [J].J Syst Evol, 2017, 55 (4): 259-290. |
| [6] | Кристина Сальмери, Кристиан Брюлло, Сальваторе Брюлло, Джампьетро Джуссо Дель Гальдо, Иван Иванович Мойсиенко. Что такое Allium paniculatum ? Установление таксономических и молекулярно-филогенетических отношений в пределах A . секта Codonoprasum [J]. J Syst Evol, 2016, 54 (2): 123-135. |
| [7] | Кумар Винод Чхотупури Госави, Шриранг Р. Ядав, К. Правин Карант, Сиддхартхан Сурвесваран. Молекулярная филогения Glyphochloa (Poaceae, Panicoideae), эндемичного рода трав из Западных Гатов, Индия. [J]. J Syst Evol, 2016, 54 (2): 162-174. |
| [8] | Юань Сюй, Чи-Мин Ху, Ган Хао.Морфология пыльцы Androsace (Primulaceae) и ее систематические последствия [J]. J Syst Evol, 2016, 54 (1): 48-64. |
| [9] | Стефани М. Икерт-Бонд, Сюзанна С. Реннер. Gnetales: последние сведения об их морфологии, репродуктивной биологии, количестве хромосом, биогеографии и времени расхождения [J].J Syst Evol, 2016, 54 (1): 1-16. |
| [10] | Ли-Э Ян, Ханг Сун, Фридрих Эрендорфер, Зе-Лонг Ни. Молекулярная филогения китайского Rubia (Rubiaceae: Rubieae) на основе ядерных и пластидных последовательностей ДНК [J]. J Syst Evol, 2016, 54 (1): 37-47. |
| [11] | Роберт Дж.Соренг, Пол М. Петерсон, Константин Ромащенко, Геррит Давидсе, Фернандо О. Сулоага, Эммет Дж. Джудзевич, Тарцисо С. Филгейрас, Джерольд И. Дэвис и Освальдо Морроне. Всемирная филогенетическая классификация Poaceae (злаковых) [J]. J Syst Evol, 2015, 53 (2): 117-137. |
| [12] | Takao KOMATSUDA. Генетика архитектуры соцветий в Hordeum [J].J Syst Evol, 2014, 52 (6): 779-782. Leave a Reply
|