
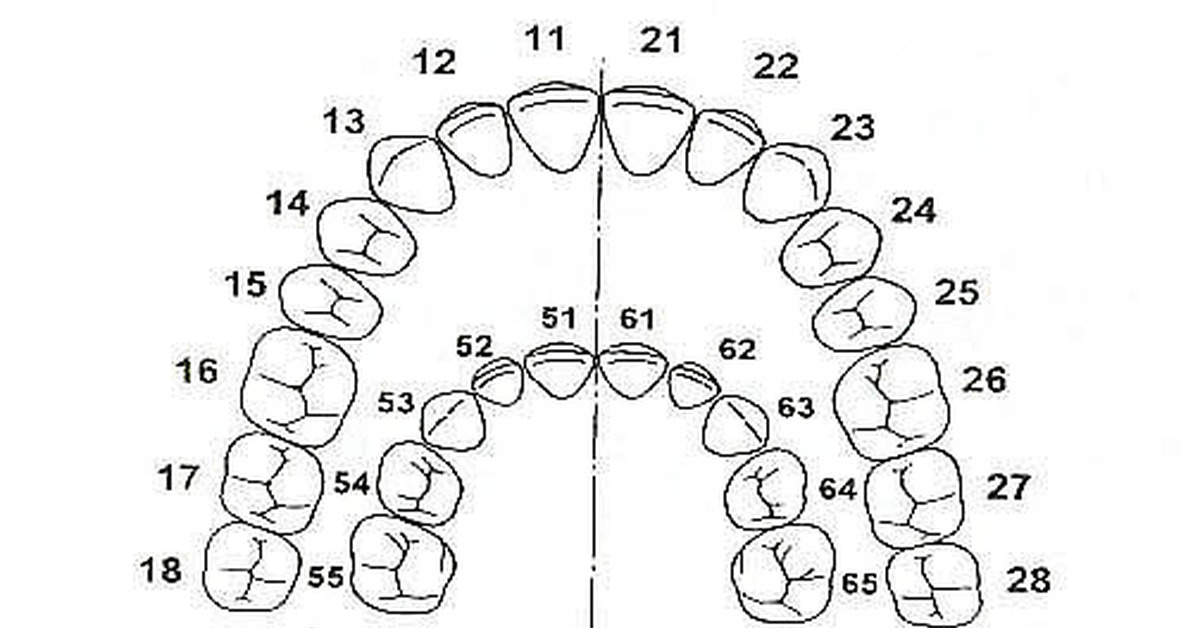
Удаление 6 зуба на верхней и нижней челюсти без боли
15.01.2020
Удаление 6 зуба затруднительно, так как он обладает большим количеством искривленных корней.
Существует несколько безусловных правил, которые станут гарантией успешного проведения данной процедуры.
Подготовка к удалению шестерки
- Правильный подбор инструментов. Существует несколько видов щипцов и элеваторов, которые могут использоваться только при удалении конкретных зубов. Ошибка в выборе инструмента приводит к достаточно неприятным последствиям: например, удаление 6 зуба верхней или нижней челюсти не может проводиться узкими щипцами — это приведет к тому, что он либо будет раскрошен, либо пациент будет чувствовать боль в момент непосредственного удаления.
- Обязательно перед процедурой нужно грамотно подобрать анестезию. В большинстве используется местная анестезия – укол лидокаина или ультракаина. Но в некоторых случаях подобные препараты не подойдут – например, если у пациента имеется индивидуальная непереносимость.

- Обследование ротовой полости перед удалением зуба – обязательная процедура. Связано такое правило с тем, что после удаления некоторых зубов могут начаться проблемы с соседними. Например, удаление шестерки подразумевает некоторое воздействие на весь ряд.
Этапы удаления
Подготовка
Стоматолог-хирург осматривает ротовую полость пациента, назначает рентген, если это необходимо. Рентгеновское обследование выявляет особенности строения, состояние и точное расположение корней, показывает наличие скрытых воспалительных процессов. Врач подбирает обезболивающий препарат и дозировку, выясняет, нет ли у пациента противопоказаний или аллергии.
Важно! Если есть страхи, сомнения или вопросы, их лучше обсудить с врачом до процедуры.

Операция
В процессе одной операции зуб надо фрагментировать по корням, а затем максимально аккуратно и малоинвазивно удалить, не разрушая окружающую шестерку костную ткань. В случае сложного удаления зуба (если шестерка скрыта десной), разрезают десну, зуб разрушают с помощью специальных инструментов и извлекают частями.
После процедуры
После удаления в лунке образуется кровяной сгусток, который защищает травмированную десну от инфекций. В течение 3 часов после операции пациенту не рекомендуется пить и принимать пищу. До полного заживления десны жевать следует на здоровой стороне рта.
простое и сложное удаление, цены в Москве
Почему удаляют зубы мудрости: подробный обзор показаний
Зачем удаляют зубы мудрости? Для этой процедуры всегда есть определенные показания, которые мы рассмотрим подробно.
Неправильное положение зуба мудрости
Если зуб мудрости дистопированный или ретинированный — его требуется удалить.
Нехватка места для прорезывания зубов мудрости
Так как зубы мудрости прорезываются позже прочих, в зубном ряду часто не остается места для их правильного прорезывания и роста. Такие зубы мудрости также подлежат удалению, потому что если их сохранить — нарушится прикус, что приведет к скучиванию и искривлению передних зубов.
Дистопированная «восьмерка» разрушает здоровый зуб
Нередко зубы мудрости прорезаются неправильно, а с наклоном и при таком положении они упираются в «семерку», в области ее шейки. Давление зуба мудрости на «семерку» приводит к постепенному разрушению эмали здорового зуба, появлению кариеса. Если диагностика выявляет подобную клиническую ситуацию — зуб мудрости также следует удалить.
Перикоронит
Коронковая часть зубов мудрости иногда оказывается покрытой слизистой оболочкой десны, которую стоматологи называют «капюшоном». Между этим капюшоном и зубом остается свободное пространство, в которое легко попадают болезнетворные организмы и вызывают воспалительный процесс, сопровождающийся образованием гноя и отеком слизистой. Так начинается перикоронит, являющийся прямым показанием для удаления зубов мудрости.
Полное разрушение коронковой части зуба мудрости
Если коронковая часть зуба мудрости разрушена так, что ее невозможно восстановить консервативным лечением — «восьмерку» следует удалить, чтобы защитить от распространения кариеса и инфекционных процессов на здоровые зубы.
Невозможность проведения качественного лечения
Зубы мудрости имеют большое количество корней сложного строения, и это затрудняет процесс терапевтического лечения «восьмерок». Если диагностика покажет, что в конкретной клинической ситуации нет возможности качественно пролечить и обработать каналы зуба мудрости, его придется удалить.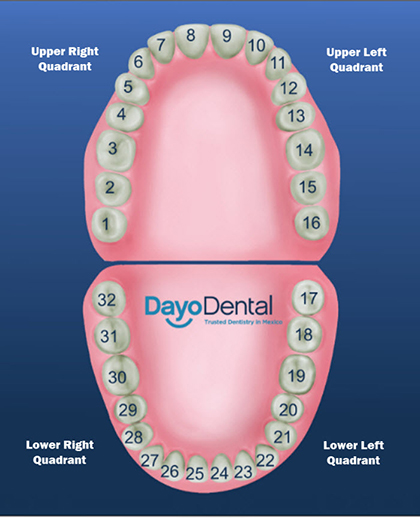
Установка брекетов
Удалять зубы мудрости иногда приходится перед установкой брекетов. Процедура удаления зубов мудрости помогает получить свободное место в ряду, для смещения других зубов, во время которого они примут правильное положение. Перед установкой брекетов именно зубы мудрости удаляют чаще всего. Почему? Все достаточно просто: при наличии всех прочих зубов, «восьмерки» не принимают участия в процессе пережевывания пищи, их удаление не повлечет за собой нарушения жевательной функции.
Нужно ли удалять зубы мудрости, если нет прямых показаний?
Еще раз обращаем ваше внимание на то, что без наличия прямых показаний зубы мудрости удалять не нужно и более того: есть ситуации, в которых необходимо сделать все, чтобы сохранить «восьмерки».
К таким ситуациям относятся:
- Необходимость в протезировании. Если 6-7 зуб удалены, потребуется опора под протез, которой вполне способен выступить здоровый зуб мудрости. Кстати, если удален 7-й зуб, зуб мудрости сдвинется и займет его место;
- Хорошее состояние зуба мудрости. Если зуб мудрости прорезался правильно, не имеет аномалий развития, у него есть зуб антагонист, он принимает полноценное участие в процессе жевания, то следует провести все необходимые мероприятия, чтобы сохранить «восьмерку».
 Кстати, если удален 7-й зуб, зуб мудрости сдвинется и займет его место;
Кстати, если удален 7-й зуб, зуб мудрости сдвинется и займет его место;Удалять ли зубы в вашем случае? На этот вопрос нельзя ответить заочно. Обязательно необходима диагностика и консультация у стоматолога. Получить профессиональный совет от специалистов по удалению зубов мудрости вы можете в нашей стоматологии в Москве, Firadent! Приходите на прием в нашу клинику: мы находимся между станциями метро Пролетарская и Волгоградский проспект.
Как удаляют зубы мудрости: простое и сложное удаление
Удаление зубов мудрости бывает простым и сложным. Решение о том, как именно будет удаляться зуб мудрости, принимается на основании области его расположения (верхняя или нижняя челюсть), особенностей его строения (ретинированный или дистопированный).
Как правило, удаление верхних зубов мудрости — простое. Почему? Потому что костная ткань в области верхней челюсти мягкая и рыхлая, поэтому хирург может вытащить третий моляр и не обломать при этом его корни. Но если перелом корня в момент извлечения зуба все же случится — процесс удаления тут же осложняется, потому, что в области верхней челюсти нет хорошего визуального доступа и требуется максимальная осторожность при обращении с хирургическим инструментом, иначе не исключены опасные травмы.
Удаление зубов мудрости на нижней челюсти в подавляющем большинстве случаев — сложное, так как кость здесь плотная, а корни у нижних «восьмерок» массивные или сильно искривленные. Поэтому почти нереально удалить зуб мудрости быстро и сразу: корни чаще всего обламываются и приходиться их вытаскивать уже после извлечения зуба из лунки. Избежать обламывания корней помогает тщательная диагностика перед удалением зуба мудрости, подразумевающая обязательный рентгеновский снимок, который покажет строение корней, их расположение в кости.
Если по снимку делаются выводы, что существуют высокие риски перелома корней зуба мудрости, его удаление проводят так: сначала, с помощью бормашины, хирург распилит зуб на несколько частей и затем, по отдельности, удалит каждую часть с корнями.
Простое удаление зуба мудрости
Выполняется оно в несколько этапов:
- Пациенту делают укол местного анестетика, подбираемого в индивидуальном порядке;
- Когда анестезия подействует, хирург специальным инструментом отделит десну от зуба мудрости;
- На зуб мудрости накладываются щипцы, которыми хирург раскачает зуб;
- Когда зуб достаточно расшатается, врач удалит его, вынув из лунки.
После удаления зуба мудрости из лунки, рана промывается специальными антисептическими растворами, в нее могут закладываться препараты, ускоряющие процессы регенерации и восстановления тканей.
Сложное удаление зубов мудрости
К сложному удалению зубов мудрости есть определенные показания.
Оно проводится в следующих случаях:
- Зуб мудрости не прорезался;
- У «восьмерки» большое количество искривленных толстых корней;
- Коронковая часть зуба мудрости разрушена так, что ее не получится зацепить щипцами;
- Третий моляр имеет сильный наклон, горизонтальное положение.
При таких обстоятельствах сразу же планируется сложное удаление зуба мудрости, и оно проводится с обязательной предварительной рентгенодиагностикой!
Сложное удаление зубов мудрости — это многоэтапная операция, подразумевающая:
- Выполнение разреза десны или распиливание коронковой части зуба мудрости на несколько частей;
- Рассверливание кости челюсти для облегчения расшатывания и извлечения «восьмерки» из лунки;
- Очищение лунки от мелких фрагментов зуба и корней, проводимое кюретажной ложкой. В отличие от обычного удаления зубов, в лунку извлеченного зуба мудрости никогда не закладываются биоматериалы. Она сама заполняется кровяным сгустком и этого достаточно для ее естественного заживления.
 В отличие от обычного удаления зубов, в лунку извлеченного зуба мудрости никогда не закладываются биоматериалы. Она сама заполняется кровяным сгустком и этого достаточно для ее естественного заживления.
В отличие от обычного удаления зубов, в лунку извлеченного зуба мудрости никогда не закладываются биоматериалы. Она сама заполняется кровяным сгустком и этого достаточно для ее естественного заживления.- Наложение нескольких швов после завершения всех необходимых манипуляций. Швы накладываются саморассасывающиеся, это значит, что для их снятия в клинику приходить не придется. Однако контрольный осмотр через пару дней после операции будет обязательным: он позволит исключить риски развития воспалительных процессов!
Операция по сложному удалению зубов мудрости длится дольше, чем простая экстракция, поэтому нередко ее проводят под общим наркозом или седацией.
Больно ли удалять зубы мудрости?
Удаление любых зубов — болезненная процедура, всегда проводимая под анестезией. Зубы мудрости не являются исключением, особенно с учетом того, что корней у них много, поэтому в случае простого удаления вам предложат местную анестезию, при необходимости проведения сложного удаления «восьмерки» — общий наркоз или седацию.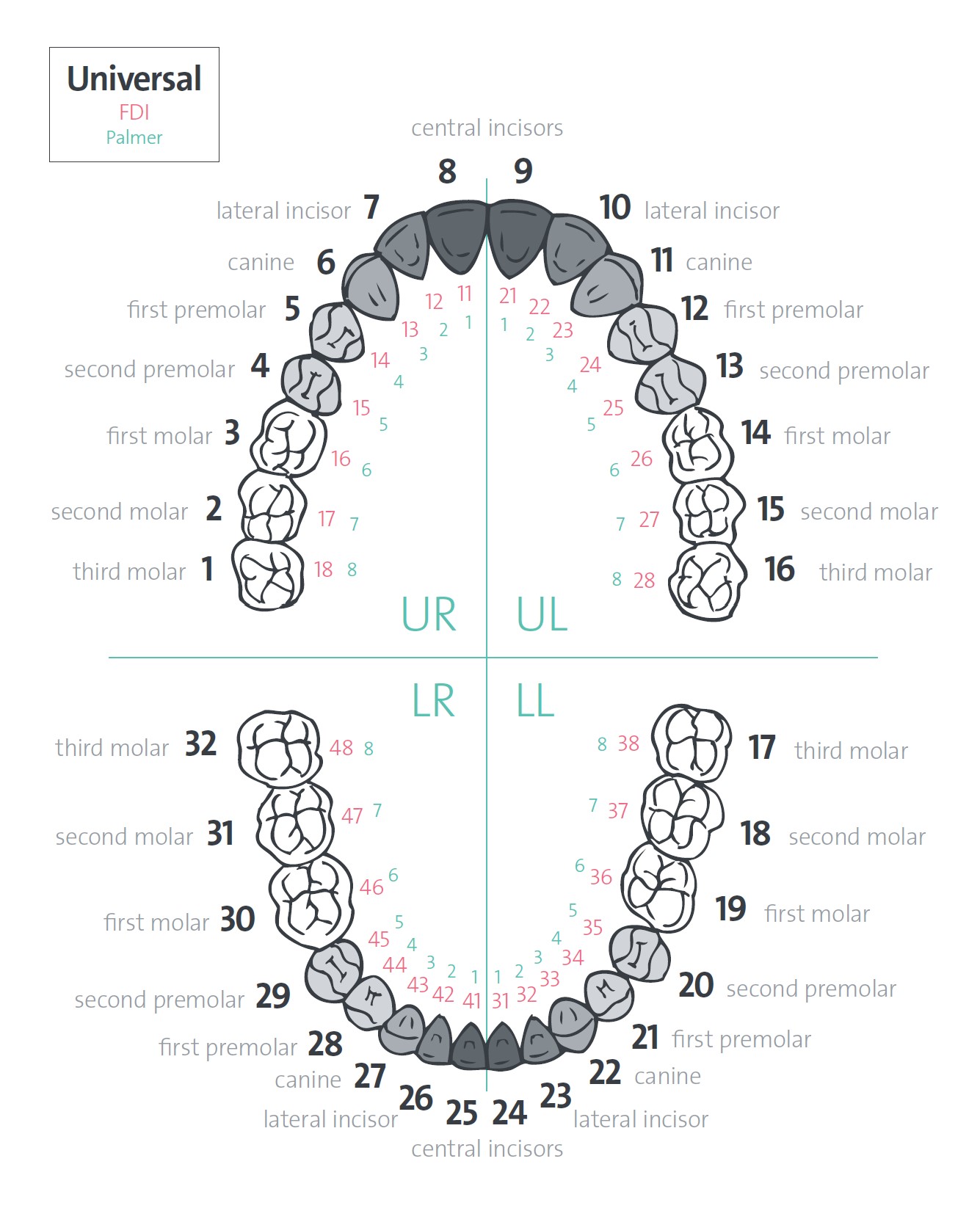
Применение общего наркоза или седации помогает провести процедуру с минимальным дискомфортом для пациента. Вы будете спать и не ощутите боли, страха или стресса. Выведение пациента из седации или наркоза осуществляется после полного завершения всех манипуляций по удалению зуба мудрости.
Что выбрать: общий наркоз или седацию?
И седация и общий наркоз могут быть использованы при удалении зубов мудрости по желанию пациента и при условии отсутствия противопоказаний. Учтите, что к общему наркозу их достаточно много, а потому придется проходить комплексное обследование, которое исключит возможные ограничения. Если вам такого обследования в клинике не предлагают — откажитесь от мысли удалять в такой стоматологии зубы под общим наркозом. Игнорирование противопоказаний при применении общего наркоза опасно для вашей жизни и здоровья!
Общий наркоз при удалении зубов мудрости чаще всего применяется тогда, когда предстоит длительная операция. Он сопровождается побочными эффектами, требует долгого и правильного выведения пациента из состояния наркоза, которое должно проходить под контролем специалистов. Поэтому, если вы решите удалять зубы мудрости под общим наркозом — внимательно отнеситесь к выбору клиники.
Поэтому, если вы решите удалять зубы мудрости под общим наркозом — внимательно отнеситесь к выбору клиники.
Известно, что сейчас услугу лечения зубов под общим наркозом предлагают пациентам многие клиники, однако по факту далеко не у всех стоматологий есть право на проведение подобных процедур и соответствующая лицензия.
У нашей клиники Firadent есть не только лицензия государственного образца на использование наркоза при стоматологических операциях, но и современное оборудование для контроля всех знаковых показателей состояния пациента при седации или общем наркозе, а в штате стоматологии работают специалисты — анестезиолог и реаниматолог.
Также мы предлагаем вам услугу удаления зубов мудрости под седацией. Седация — термин, обозначающий состояние легкого медикаментозного сна, при введении в который пациент остается в сознании и даже способен выполнять указания стоматолога. Седация избавляет от болевых ощущений, от стресса и страхов, которые мешают проведению качественного лечения и способны привести к психологической травме. Медикаментозный сон не сопровождается побочными эффектами, не требует длительного выхода под контролем врачей, после того, как человек проснется — он может покинуть клинику и отправиться по своим делам.
Медикаментозный сон не сопровождается побочными эффектами, не требует длительного выхода под контролем врачей, после того, как человек проснется — он может покинуть клинику и отправиться по своим делам.
Что выбрать в вашем случае — удаление зубов мудрости под общим наркозом или седацией? Запишитесь на консультацию в Firadent и врачи клиники подберут для вас оптимальный вариант анестезии для удаления зубов мудрости.
7 проблем зубов мудрости
Зуб мудрости — третий моляр, 8-й зуб в ряду (его еще называют «восьмерка»), период его прорезывания 14-25 лет. Внешне он ничем не отличается от остальных, однако особенности его развития и роста могут приводить к воспалительному процессу и к различным заболеваниям.
Проблема 1 — ретинированный зуб мудрости! Представляет собой полностью сформировавшийся, но оставшийся в челюсти либо частично прорезавшийся зуб. Существует два варианта развития событий. Первый – зуб совсем не будет беспокоить своего хозяина, второй – со временем такой зуб начнет беспокоить, а так же двигать соседние зубы в процессе прорезывания, что ведет к изменению прикуса. Если вовремя не удалить такой зуб, то впоследствии могут возникнуть проблемы со здоровыми зубами.
Если вовремя не удалить такой зуб, то впоследствии могут возникнуть проблемы со здоровыми зубами.
Проблема 2 — дистопированный зуб мудрости! Это зуб, рост которого происходит с отклонениями от нормы (под неправильным углом, не той стороной). К сожалению, в большинстве случаев дистопированный зуб мудрости удаляют. Связано это с тем, что неправильно расположенные «восьмерки» травмируют окружающие ткани, провоцируя возникновение воспалений и абсцессов, а также негативно влияют на соседние зубы и прикус в целом.
Проблема 3 — затрудненное прорезывание зуба мудрости. Так как на месте прорезывания зуба мудрости не было молочного зуба, то десна в том месте не приспособлена к прорезыванию. Из-за этого часто наблюдается набухание места прорезывания зуба мудрости и порой невыносимые боли в десне, а иногда в ухе и даже в шее. В некоторых случаях боль настолько невыносима, что человек с трудом может открыть рот. В таких ситуациях необходимо обратиться за помощью к специалисту, который поможет преодолеть боль.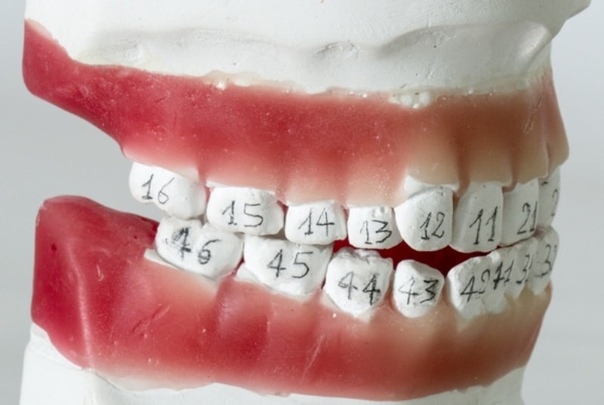 В среднем, зуб мудрости может прорезываться от нескольких месяцев до нескольких лет.
В среднем, зуб мудрости может прорезываться от нескольких месяцев до нескольких лет.
Проблема 4 — кариес «восьмерки». Кариес — пожалуй, самое частое заболевание полости рта. Ему подвержены любые зубы, но восьмерки — в особенности. Причиной этого заболевания является скопление бактериального налета и плохая гигиена. Несмотря на нерациональность сохранения восьмерки, иногда врач рекомендует отказаться от крайних мер и назначает лечение восьмого зуба. Но более распространенным результатом после постановки такого диагноза является удаление пораженного кариесом зуба.
Проблема 5 — кариес корня зуба мудрости! Главное отличие корневого кариеса от всех остальных — это его локализация глубоко под десной. И если поражение в пришеечной и прикорневой части зуба можно обнаружить при визуальном осмотре, то кариес корня незаметен и потому наиболее опасен из всех. Кариес может развиться на корне «восьмерки» из-за ее неправильного прорезывания, когда корень оголен, а так же есть опасность развития кариеса корня на соседнем 7-ом зубе, в случае, когда зуб мудрости в стадии прорезывания двигает соседний зуб, оголяя его корень, являясь препятствием для нормальной гигиены в этой области.
Проблема 6 — перикоронарит! В процессе прорезывания зуба мудрости над ним появляется десневой капюшон, который может воспалиться, также он доставляет невыносимую боль. В этом месте происходит гнойное воспаление. Если вовремя не провести процедуру по удалению десневого капюшона, то воспаление может перекинуться на надкостницу и даже кость.
Проблема 7 — периодонтит! Хронический периодонтит зуба мудрости редко беспокоит больного, протекает практически бессимптомно. Проявляются незначительные боли только при накусывании и постукивании. На десне в проекции зуба мудрости может существовать свищ, из которого будет выделяться скудное гнойное отделяемое. Острый периодонтит зуба мудрости проявляется возникновением резкой боли при накусывании, на фоне постоянных болей ноющего характера, наблюдается небольшая подвижность зуба, может появится припухлость десны/щеки.
В любой форме и стадии болезни, если зуб мудрости сильно беспокоит, это серьезный повод для обращения к стоматологу.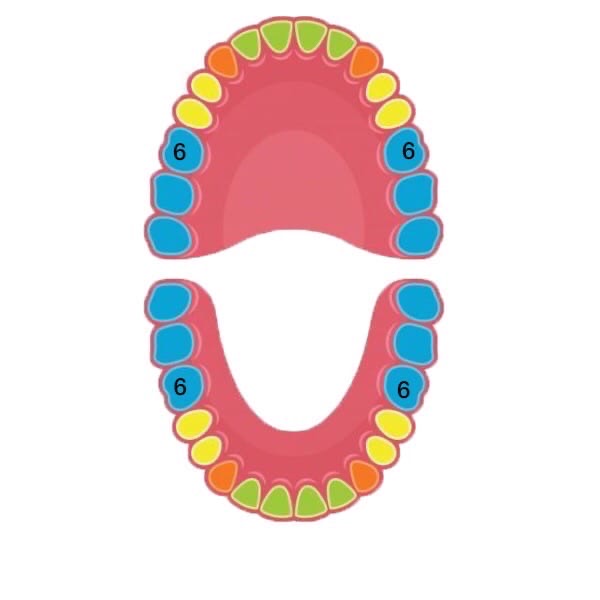 Максимально эффективным будет обращение на ранней стадии развития патологии. Поэтому рекомендуется не затягивать с регулярным профилактическим осмотром, не дожидаясь, пока даже легкое воспаление, перейдет в более осложненные формы.
Максимально эффективным будет обращение на ранней стадии развития патологии. Поэтому рекомендуется не затягивать с регулярным профилактическим осмотром, не дожидаясь, пока даже легкое воспаление, перейдет в более осложненные формы.
Держись за зуб! Московский комсомолец
Ректор МГМСУ Николай Ющук: “Среди стоматологов миллионеров нет”
С детства я боюсь зубных врачей: визг бормашины приводит в ужас, от медицинских запахов подгибаются ноги. Такая проблема есть не только у меня одного: около 50% людей обращаются к стоматологам лишь в крайнем случае, а 20% предпочитают терпеть до последнего. Однако современные методики позволяют проводить лечение почти без боли. Уже на студенческой скамье будущих врачей учат щадящим технологиям. О новых направлениях в подготовке студентов рассказал ректор Московского государственного медико-стоматологического университета академик РАМН профессор Николай ЮЩУК.
МГМСУ сегодня: 9 факультетов, 105 кафедр, более 10 тысяч студентов, 3260 преподавателей и служащих, в т.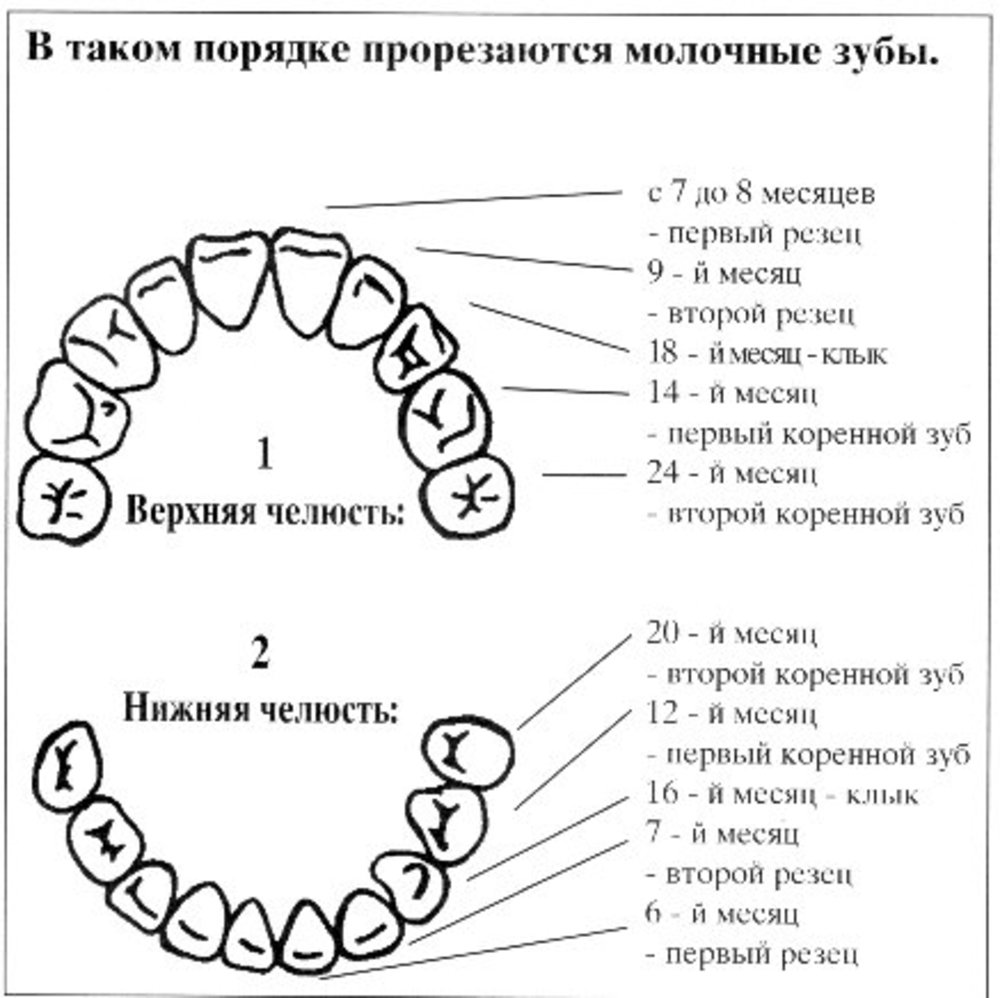 ч. 700 профессоров и доцентов, 20 академиков и членов-корреспондентов РАН и РАМН.
ч. 700 профессоров и доцентов, 20 академиков и членов-корреспондентов РАН и РАМН.
— Основатель стоматологического дела в России и первый ректор вуза профессор Александр Евдокимов любил повторять: “Держитесь за зуб, не пропадете!” Это верно: наши выпускники нарасхват. А все потому, что ребят учат специалисты с мировым именем. Например, профессор Гарри Барер, возглавляющий кафедру госпитальной терапевтической стоматологии, академик Леонид Персин, возглавляющий одну из лучших в Европе клиник ортодонтии, профессор Рабинович, один из родоначальников борьбы с болью при лечении зубов. Благодаря разработкам его кафедры можно помогать людям, страдающим и гипертонией, и остеопорозом, и аллергией, и сахарным диабетом. Профессор, декан факультета Сергей Иванов — разработчик отечественных имплантологических систем, великолепный хирург. На кафедре детской хирургической стоматологии профессора Светланы Дьяковой оперируют детей со сложными патологиями челюстно-лицевой области: опухолями и врожденными уродствами. В МГМСУ работают такие светила отечественной медицины, как Валерий Шумаков, Лейла Адамян, Лео Бокерия, Дмитрий Пушкарь, Владимир Крылов, Анатолий Мартынов, Евгений Соколов, Игорь Лебеденко, Магомед Дебиров…
В МГМСУ работают такие светила отечественной медицины, как Валерий Шумаков, Лейла Адамян, Лео Бокерия, Дмитрий Пушкарь, Владимир Крылов, Анатолий Мартынов, Евгений Соколов, Игорь Лебеденко, Магомед Дебиров…
— “МК” уже сообщал, что университет открыл кафедры в необычном месте — в “Матросской Тишине”…
— Да, зарождается новое направление — подготовка специалистов пенитенциарной медицины. А самый большой конкурс у нас — 6,5 человека на место — на факультете клинической психологии. Во время терактов и катастроф психологи нужны не меньше, чем хирурги. Их работа — помощь людям в стрессовых ситуациях.
Новый факультет — экономики здравоохранения. На нем готовят заместителей главврачей по экономике и организаторов здравоохранения. Это менеджеры от медицины. Раньше считалось, что “широкий” экономист может работать в любой сфере, но в медицине своя специфика. Поэтому на факультете обучают основам врачебного дела и тонкостям экономики. Популярен факультет среднего медобразования, где учат на зубных техников и стоматологических гигиенистов. В этом году при поддержке губернатора Бориса Громова в МГМСУ открылся факультет по подготовке специалистов для Московской области. Приняли 25 человек на стоматологию и 65 — на лечебное дело. Ребята учатся за счет областного бюджета, а потом три года отрабатывают в Подмосковье.
— Стоматология всегда считалась прибыльным делом… Многие шли учиться, надеясь потом хорошо зарабатывать. Нет ли сегодня перекоса в пользу платного обучения за счет бюджетного?
— В этом году мы приняли почти 1000 человек на бесплатные места и всего 300 — на коммерческие. Требования ко всем одинаковые: не можешь учиться — отчислим. Университет дорожит своей репутацией. Пусть лучше человек найдет себя в другой области, чем станет плохим врачом. Ведь медицина — это не только хорошие заработки, но и адский труд. У наших врачей есть хорошая поговорка: среди стоматологов миллионеров нет. Стоматологи, кстати, входят в первую группу риска — велика опасность заразиться гепатитом или ВИЧ-инфекцией и другими заболеваниями, передающимися с кровью, слюной и воздушно-капельным путем. Пациент, когда садится в кресло, не всегда говорит, чем болен…
— Сейчас много говорят о двухуровневой системе высшего образования: бакалавр—магистр…
— В медицине не может быть бакалавров. Четыре года — слишком мало для подготовки врача, нужно не меньше пяти. Реформировать отечественное образование следует, но его фундаментальную основу необходимо сохранить.
— Через несколько лет принимать во все российские вузы будут только по результатам Единого госэкзамена…
— По тестам нельзя определить, подходит ли абитуриенту профессия врача. Отличные ЕГЭ-оценки еще не означают, что школьник может стать хорошим терапевтом или стоматологом. Нужен талант, любовь к людям, терпение, сострадание… ЕГЭ — это эксперимент, который вряд ли подходит медвузам в нынешней форме.
— В университете всегда было много иностранцев. Не боятся ли они сейчас приезжать в Россию?
— У нас учится более 500 студентов из 64 стран. В основном из дальнего зарубежья: Индия, Китай, Эквадор, Пакистан, Кипр, Греция… Есть даже ребята из США, Германии, Японии и Израиля. Их расчет понятен: при хорошем качестве образования цена знаний в России ниже, чем в большинстве западных стран.
— Вопрос, волнующий многих абитуриентов: судьба военной кафедры. Министерство обороны обещает сократить подготовку офицеров запаса…
— Военные врачи нужны всегда, особенно специалисты в области челюстно-лицевой хирургии. Надеемся, что Минобороны примет правильное решение.
Материал: Игорь Сергеев
показания, трудности, особенности удаления воьмерки
31.05.2018
Зубы мудрости вырастают нормально только в 20% случаев, а 8 из 10 пациентам их нужно удалять еще во время прорезывания. Даже если они заняли физиологическое положение во рту, на них часто развивается кариес и пульпит. При этом лечение осложнено затрудненным доступом и запутанным строением корневой системы.
Происхождение и роль «восьмерок» в зубочелюстной системе
Проблемы с прорезыванием зубов мудрости заключаются в:
- недостаточном пространстве на челюсти;
- отсутствием предшественников – молочных зубов;
- времени появления – третьи моляры вырастают в 18 – 25 лет, когда костные ткани уже полностью сформированы и уплотнены.
Даже нормальный рост сопровождается неприятными симптомами: набуханием десны, болью, зудом, кровоточивостью, субфебрильной температурой (до 37,5°С). Если же прорезывание осложненное, эти симптомы усиливаются, развивается воспаление слизистой, подчелюстных лимфоузлов.
Сложности обусловлены происхождением 3-их моляров. Они – яркий пример атавизма. Предкам человека приходилось пережевывать пищу плохо или вовсе не обработанную термически. Была нужна большая площадь для первичной обработки еды. Со временем продукты стали мягче, а пережевывать их – проще. Поэтому необходимость в дополнительных зубах отпала. В ходе эволюции зубочелюстная дуга уменьшилась на 10 – 12 мм, для «восьмерок» не осталось места.
Зубы мудрости – рудиментарный орган. Они почти не участвуют в измельчении пищи. С этой функцией справляются 1-е и 2-е моляры – «шестерки» и «семерки». Однако не стоит спешить избавляться от них. Иногда они могут пригодиться.
Их стараются сохранить, когда были удалены «семерки» и соседние «шестерки» или высока вероятность потерять их в ближайшее время. В этом случае их используют как опору для протезирования зубов.
Но к таким мерам прибегают редко. У последних элементов слабые корни, они с трудом выдерживают ту малую нагрузку, которая приходится на них и не удержат протез долго. Также из-за отдаленного расположения за ними сложно ухаживать. Будет постоянно скапливаться налет, который приведет к отложению камня, гиперемии десен и потере «моста» через пару лет. Поэтому лучше решение – установить имплантат.
Возможные трудности с зубами мудрости
Проблемы с третьими молярами возникают еще на этапе их прорезывания. К ним относят:
- Дистопию – неправильное положение зубов. Аномалийное положение сопровождается травмами слизистой, что может привести к развитию патологических изменений эпителия, в том числе злокачественных; упираясь в соседние, интактные зубы, «восьмёрки» провоцирует деформацию зубной дуги.
- Ретенцию – полное или частичное непрорезывание. Полуретенированные зубы обязательно удаляют, так как они приводят к травмам и воспалению десен, смещению интактных коронок.
- Перикоронарит – воспаление наддесневого капюшона. Заболевание развивается из-за скопления остатков пищи и бактериального налета в свободном пространстве под десной. Вычистить их щеткой невозможно. Постепенно развивается покраснение и отечность, образуется жидкость с гнойным содержимым (экссудат), возникает болезненность, нарушается симметричность лица, повышается температура, увеличиваются региональные лимфатические узлы.
Даже если «восьмерки» нормально прорезались, они часто подвергаются кариозным поражениям. Связано это с их отдаленным расположением и, как следствие, затрудненной гигиеной.
Полностью убрать налет сложно. Скапливаясь, отложения приводят к поражению эмали и дентина. Если не устранить дефект на этом этапе, он перетечет в пульпит (поражение сосудисто-нервного пучка), а затем – в периодонтит (инфекционное поражение верхушки зуба). Положение может осложняться гингивитом (воспалением десен), формированием гранулем и кист на корнях – капсул с гноем.
Показания для лечения и удаления
Решение о сохранении или удалении принимает только врач, основываясь на клинической картине и данных рентгенологической диагностики. Зуб оставляют, если:
- он нормально прорезался и не мешает другим единицам;
- у него есть антагонист – еще одна «восьмерка» с той же стороны на противоположной челюсти;
- он устойчив, играет роль «сдерживающего» фактора – предотвращает смещение соседних коронок.
Лечение зуба мудрости мало отличается от терапии других зубов. При кариесе также убирают поврежденные ткани, обеззараживают полость, устанавливают пломбу. Единственное ограничение – материал для восстановления. Предпочитают использовать стеклоиономерный цемент, а не светоотверждаемый полимер. Так как щека находится близко к коронке, невозможно полноценно высушить полость и подвести полимеризационную лампу к обрабатываемому участку.
Иначе выглядит ситуация при пульпите. В этом случае большинство стоматологов склоняются к экстракции. У 3-их моляров запутанная корневая система, непредсказуемое количество корневых каналов, ложных и микроканальцев. Из-за осложненного доступа трудно работать эндодонтическими инструментами. Даже с дентальным микроскопом полноценно очистить и запломбировать каналы почти невозможно.
Врач клиники обязательно консультирует пациента о возможных осложнениях.
Удаление обязательно проводят при:
- осложненном прорезывании;
- дистопии и ретенции;
- нехватке места;
- невозможности терапевтического лечения;
- периодонтите, гранулемах, кистах;
- рецидивирующем перикороните;
- разрушении соседней «семерки»;
- угрозе искривления прикуса.
Еще одно показание для удаления – планируемое выравнивание прикуса. Для этого необходим запас пространства на челюсти, куда будут передвигаться скученные единицы. А если «восьмерки» начнут расти уже после исправления прикуса, они исказят ее повторно.
Особенности удаления зубов мудрости и реабилитация
Так как экстирпация сопровождается высокой травматичностью, невозможно избежать некоторых осложнений. Они считаются нормальными и проходят самостоятельно. К ним относят:
- болит десна после удаления зуба мудрости;
- отек слизистой;
- незначительное посинение десны;
- незначительное увеличение температуры.
Иногда развиваются серьезные последствия:
- выпадение кровяного сгустка из лунки – он нужен, чтобы предотвратить попадание пищи и патогенных микроорганизмов в рану;
- альвеолит или «сухая лунка» – инфицирование полости, происходит из-за вымывания сгустка;
- длительное первичное или вторичное кровотечение – появляется при обширных повреждениях тканей, его развитию способствуют сахарный диабет, плохая свертываемость крови, прием кроворазжижающих препаратов (аспирина и др.).
Чтобы избежать патологических осложнений, необходимо четко следовать указаниям врача стоматологии. Для предотвращения рекомендуют:
- наложить швы: даже если рана небольшая, ее лучше ушить – это снизит риск последствий на 90%;
- принимать антибиотики – обычно назначают «Амоксициллин», «Линкомицин», «Амоксиклав», их выписывают обязательно, если экстирпация проводилась на фоне инфекционного процесса;
- первые часы после экстракции не есть и не пить;
- придерживаться диеты несколько дней – исключить горячую, холодную пищу, напитки и продукты, которые раздражают слизистую;
- не греть прооперированный участок;
- чтобы снять отек, можно прикладывать к щеке холодные компрессы на 5 – 10 минут;
- снять боль можно анальгетиками и противовоспалительными препаратами – «Кетанов», «Нимесил» и др.;
- тщательно поддерживать гигиену ротовой полости;
- использовать антисептические растворы для полоскания – хлоргексидин, «Мирамистин», «Стоматофит», первые 2 – 3 суток делают ротовые ванночки, чтобы не вымыть сгусток.
Если удаление зуба мудрости проводил квалифицированный стоматолог, а пациент четко придерживался его советов, заживление пройдет быстро и без последствий. Уже через пару дней лунка начнет затягиваться, а спустя несколько недель от раны не останется следа.
лечить или удалять. Нужно ли удалять зуб мудрости, цены
В стоматологической практике достаточно распространенной является дилемма — зубы мудрости — лечить или удалять. Эти зубы, по сути, не выполняют никаких функций, находятся в труднодоступный местах, они имеют глубокие корни, поэтому проводить лечение достаточно сложно. При таких показаниях, казалось бы, ответ на вопрос: зуб мудрости — лечить или удалять, все аргументы в сторону удаления. Но при этом, если размышлять на перспективу, то 8-ой зуб может стать надежной опорой для установки мостовидного протеза. Стоит сделать ударение на том, что решать этот вопрос, есть смысл только, если расположение зуба анатомически правильное. Если вырос горизонтально 8 зуб, лечить или удалять — все показания к удалению.
Показания к удалению зуба мудрости
Для того, чтобы определиться с собственной стратегией касательно лечения или удаления 8-го зуба, Вам нужно обратиться к специалисту. Он направит Вас на рентгеновский снимок, осмотрит зуб и на базе этой информации расскажет об анатомических особенностях Вашего клинического случая. Если стоматолог выделит показания к удалению зуба мудрости, то такой зуб лучше не сохранять, иначе цена за его лечение себя не оправдает.
Рассмотрим более детально, когда нужно удалять зуб, какие показания к удалению зуба мудрости существуют:
- Неправильный рост зуба — 8-й зуб очень часто растет не вертикально, а горизонтально. С уклоном к щеке или к языку. Такой зуб не участвует в выполнении функции жевания, зато постоянно травмирует мягкие ткани ротовой полости.
- Появление скученности зубов, когда начал прорезаться 8-й зуб.
- Сильный кариес, который разрушает коронку зуба.
- Разрушение 7-го коренного зуба из-за нарушения расположения 8-го.
В любом случае пациент сам принимает решение относительно того нужно ему лечение или удаление зуба мудрости.
Нужно ли удалять зуб мудрости, если он не болит
Большинство людей сталкиваются с проблемным ростом 8-го зуба, но есть и те, которые не знают о дискомфорте, когда есть 8-й зуб. Поэтому, нужно ли удалять зубы мудрости, если они не болят? Для того, чтобы перестраховаться от многих неприятных ситуаций в будущем и не платить лишний раз цену за стоматологическое лечение, пациенту лучше сделать рентген и проконсультироваться с доктором. Если зубы не болят, не поражены кариесом и рентгеновский снимок показывает, что они никоим образом не влияют на здоровье остального зубного ряда, то их можно оставить. Возможно в старости, в случае потери жевательных зубов, они частично возьмут на себя жевательную функцию. Нужно ли удалять зуб мудрости — зависит от каждого индивидуального клинического случая, иногда лечение является более рациональным решением.
Когда нужно непременно удалять зуб мудрости
Есть показания, когда нужно удалять зуб мудрости обязательно. Например, гнойный процесс, который находится в запущенной стадии. Если сломалась коронка зуба, при этом ее нельзя запломбировать или применить ортопедическое лечение. Осевой перелом зуба — также является показанием, когда нужно удалять зуб мудрости.
Зуб мудрости, цены на удаление
При наличии показаний на удаление зуба мудрости, цена на услуги хирурга-стоматолога может существенно отличаться. В первую очередь, она зависит от сложности случая. Если корни, которые имеет зуб мудрости глубокие — удаление будет сложным, цена, соответственно также будет выше. Для того, чтобы вырвать зуб, стоматолог применяет местную анестезию, бывают ситуации когда требуются дополнительные надрезы и применение специальных инструментов, чтобы извлечь моляр. Ориентировочная цена на удаление зуба мудрости представлена на сайте компании «ДАНТИСТ&КО». Но окончательная стоимость процедуры может быть сформирована только после завершения процедуры.
Безболезненное лечение зубов | ООО «Медсервис»
Прогресс в развитии технологий лечения зубов дошёл до такого уровня, что находясь в кресле у стоматолога вы можете ощущать себя посетителем салона красоты. Во-первых, лечение действительно проходит безболезненно, а во-вторых, те методики, которые стали доступны и применяются в нашей клинике, позволяют проводить реставрацию зубов любой степени сложности.
Сегодня, лечение разрушенного зуба это уже не столько постановка обычной пломбы, сколько высоко эстетичная реставрация зуба, позволяющая восстановить анатомию зуба.
Большинство людей сегодня уже знают: стоматология — это не больно. Специалисты применяют уникальные анестезирующие и седативные препараты, которые обеспечивают комфортное лечение и не оказывают вредного воздействия на организм. Мы расскажем о седации: в чем ее преимущества и чем она отличается от традиционного наркоза.
Седация — это поверхностный сон, который предполагает возможность в любой момент разбудить пациента. Комплекс мер, нацеленных на полное расслабление, обеспечивают успокоение пациента на все время проведения стоматологических манипуляций. Седация в стоматологии проводится при помощи специальных седативных средств и вместе с традиционной анестезией является прекрасной альтернативой наркозу
Основные отличия седации от общего наркоза заключаются в том, что:
- Не происходит блокировки глотательных, кашлевых и других рефлексов. Это обеспечивает абсолютную безопасность стоматологического лечения.
- Пациент находится в сознании, то есть врач может полностью контролировать его состояние. Более того, из состояния поверхностного сна человека очень легко вывести.
- Предыдущий пункт определяет еще одно важное преимущество седации: пациент, находясь в расслабленном состоянии, может общаться с врачом.
Противопоказаний к седации практически не существует. Ее широко применяют при лечении детей и взрослых (но предварительная консультация все-таки необходима).
Преимущества седации:
- Вы не испытывает никакого стресса, страха и психологического дискомфорта.
- За одно посещение стоматолога может быть проведено больше процедур: вы можете пройти одновременно лечение и имплантация зубов!
- Вы сможете провести в кресле стоматолога до 8 часов, что практически невозможно без применения седации. Уменьшается число посещений клиники .
- Никаких неприятных воспоминаний!
- Вы экономите свое время!
Если Вас заинтересовала информация о «спящей стоматологии», если Вы решили удалять, лечить или протезировать зубы под седацией, запишитесь на консультацию по телефону (3476) 320-900 и обязательно сообщите об этом Вашему врачу. Врач даст все необходимые разъяснения и согласует с Вами время и объем стоматологической помощи с применением данной технологии.
Подробнее по тел. 8-800-250-32-90.
Число зубов — обзор
Человек
Первый набор молочных или молочных (проливных) зубов номер 10 в каждой челюсти, что в сумме составляет 20. С каждой стороны, начиная спереди, есть два резца (лезвие- например, режущие зубы), один клык (толстый, пронзительный, похожий на клык) и два коренных зуба сзади (более широкие зубы с бугорками для измельчения пищи). Молочные зубы прорезываются через определенные промежутки времени по мере освобождения места в растущих челюстях. Резцы появляются первыми в возрасте примерно 6 месяцев, а последние — задние зубы (вторые коренные зубы) примерно в два с половиной года.В течение следующих 3-4 лет у младенца будет полный молочный зубной ряд из 20 зубов (рис. 9.8).
Рисунок 9.8. Нижняя челюсть ребенка 3 лет с пятью молочными зубами с каждой стороны. Задние зубы — это вторые молочные коренные зубы, а зубы по средней линии спереди — это первые (центральные) резцы.
Иногда молочные резцы прорезываются рано и присутствуют при рождении; их называют натальными зубами. Вокруг таких младенцев существует множество суеверий. У некоторых пораженных новорожденных, таких как Ганнибал, Людовик XIV и Наполеон Бонапарт, наличие натальных зубов считалось очень благоприятным признаком.В других, таких как Ричард III и Иван Грозный, это считалось признаком зла. Дети, рожденные с зубами, часто становились объектами детоубийства (которое, несомненно, продолжается в некоторых частях мира), поскольку это было связано с колдовством. Наличие лишнего пальца, как у Ивана Грозного, тоже не помогло бы их делу.
В первые 2–3 года, когда прорезываются молочные зубы, ребенок может испытывать некоторый дискомфорт. Симптомы этого «прорезывания зубов» могут быть разными. Десны могут болеть, щеки покраснеть, ребенок может быть раздражительным и иметь жар.Вплоть до восемнадцатого и девятнадцатого веков прорезывание зубов ошибочно считалось прямой причиной смерти маленьких детей. На рис. 9.9 показаны зарегистрированные случаи смерти в Лондоне за 1703 год. Из 26 720 случаев смерти 1305 указаны как «от зубов». Конечно, настоящие причины не имели ничего общего с зубами и заключались в антисанитарных и убогих условиях жизни, отсутствии подходящей пищи и инфекционных заболеваниях, особенно среди бедных.
Рисунок 9.9. Страница из Биллей о смертности приходов Лондона за 1703 год.
Источник : Предоставлено библиотекой Королевского медицинского общества, Лондон.По мере роста ребенка челюсти увеличиваются, о чем свидетельствуют промежутки между зубами. Теперь требуется большая поверхность зубов, чтобы иметь дело с увеличенным количеством и разнообразием пищи, потребляемой растущим ребенком. Это достигается двумя способами, а именно за счет замены зубов большего размера и за счет большего количества зубов. На рис. 9.10 изображен череп ребенка пяти с половиной лет, но с удаленной костью, чтобы показать основные постоянные зубы на разных стадиях развития и первые постоянные коренные зубы, которые вот-вот прорежутся за задним молочным зубом.
Рисунок 9.10. На левой стороне черепа ребенка пяти с половиной лет видны прорезавшиеся по пять молочных зубов на каждой челюсти. Кость была отсечена, чтобы показать некоторые постоянные зубы (P), развивающиеся под прорезавшимися молочными зубами, и первый постоянный коренной зуб (M), который вот-вот прорежется за последним молочным зубом.
Источник : Предоставлено Хантерианским музеем Королевского колледжа хирургов.У людей замена зубов происходит в течение примерно 12 лет, начиная с 6–18 лет.Каждый из первых трех передних молочных зубов заменяется постоянным зубом большего размера. (Два задних молочных зуба, молочные коренные зубы, заменяются двумя постоянными премолярами, которые на самом деле немного меньше по длине.) При выпадении корни молочных зубов разрушаются / рассасываются основным прорезывающимся постоянным зубом, что приводит к их расшатыванию. . Если постоянный зуб не может развиться ниже молочного зуба, молочный зуб может сохраниться во взрослой жизни.
Молочные зубы выпадают и периодически заменяются постоянными зубами, причем первые нижние постоянные резцы прорезываются первыми примерно в 6-7 лет, а вторые постоянные премоляры (или двустворчатые, что означает два бугорка) прорезываются в последнюю очередь примерно в 11-12 лет. годы.Примерно в возрасте 6–12 лет прорезавшийся зубной ряд состоит из смеси молочных и постоянных зубов, стадии «смешанного прикуса» (рис. 9.11). После своего первоначального появления постоянному зубу требуется около года, чтобы достичь необходимой высоты, чтобы прикусить его зуб-антагонист на противоположной челюсти. Хотя щели периодически появляются после выпадения молочного зуба и до прорезывания постоянного наследника, они не мешают ребенку нормально есть.
Рисунок 9.11. Нижняя челюсть ребенка 11 лет со смешанным прикусом, с молочными (D) и постоянными (P) зубами. Задний зуб представляет собой второй постоянный коренной зуб, который только что прорезывается во рту.
По мере увеличения длины каждой челюсти сзади появляется достаточно места для добавления трех больших шлифовальных зубов с каждой стороны, постоянных коренных зубов. У этих зубов нет предшественников. Эти коренные зубы добавляются периодически примерно каждые 6 лет: первые постоянные коренные зубы прорезываются в 6 лет за последним молочным зубом (см. Рисунок 9.10), вторые постоянные коренные зубы позади него примерно в 12 лет (рис. 9.11) и третьи постоянные коренные зубы (так называемые зубы мудрости, потому что к моменту их прорезывания один из них старше и мудрее) примерно в 18 лет. возраст. В итоге каждая челюсть имеет 16 зубов (по восемь с каждой стороны) (рис. 9.12), что в сумме дает 32. Полный зубной ряд взрослого человека, начиная с передней части, имеет два резца, один клык, два премоляра (заменяющие два молочных моляра) и три коренные зубы с каждой стороны.
Рисунок 9.12. Нижняя челюсть показывает полный зубной ряд взрослого, состоящий из восьми зубов в каждой половине. Поскольку это молодой человек около 25 лет, зубы мало изнашиваются, а точки контакта между соседними зубами маленькие (стрелки). Сравните с рисунком 9.15.
Когда постоянные коренные зубы впервые прорезываются с их гладкими, неизношенными закругленными бугорками, они не подходят для жевания и требуют длительного периода «притирки», чтобы немного их приплюснуть для лучшего функционирования. Вы можете себе представить, насколько неэффективно было бы, если бы постоянные коренные зубы были внезапно заменены, когда они, наконец, изношены и начали нормально функционировать, и весь процесс пришлось бы начинать заново! (Это дополнительно проиллюстрировано на рисунке 9.16.)
Сегодня у людей зубы мудрости отсутствуют у 25% населения, или, если они присутствуют в нижней челюсти, они остаются непрорезавшимися (ретинированными) еще у 25% населения из-за отсутствия зубов. пространство (рисунок 9.13). В случае частичного прорезывания может произойти застой пищи вокруг пораженного коренного зуба и привести к болезненному воспалению десен. Это основная причина удаления зубов мудрости.
Рисунок 9.13. Рентген зубного ряда показывает два ретенированных моляра нижней трети (стрелки).
Одно из объяснений широкого распространения ретенированных зубов мудрости связано с природой современной диеты во многих так называемых «развитых» странах. В «примитивных» обществах диета часто бывает жесткой, грубой и абразивной, требующей значительного пережевывания. Это приводит к быстрому износу большей части шлифовальной поверхности. Износ может быть настолько обширным, что обнажает пульпу зуба и вызывает болезненные абсцессы. Это обычно наблюдается в черепах ранних цивилизаций (рис. 9.14), где, например, хлеб содержал некоторое количество песка и других абразивных частиц во время его приготовления.Поскольку зубы очень слабо перемещаются в лунках во время жевания, также наблюдается износ в точках контакта между соседними зубами: эти контакты узкие, когда зубы впервые прорезываются из-за их естественного выпуклого контура. Можно ожидать, что из-за этого износа между зубами останутся небольшие зазоры. Однако по какому-то неизвестному механизму эти промежутки автоматически закрываются, когда все зубы медленно перемещаются вперед (мезиальный дрейф). Со временем при дальнейшем износе точки контакта между соседними зубьями становятся намного шире (сравните Рисунок 9.12 с рисунком 9.15). В таких группах, как эскимосы и австралийские аборигены, чья диета (по крайней мере, до недавнего времени) состояла из жесткой пищи, это вызывало до 3–4 мм износа в точках контакта с каждой стороны челюсти. По мере того, как зубы перемещаются вперед, чтобы заполнить промежутки, это дает достаточно места в задней части челюсти для прорезывания зубов мудрости, поэтому ретинированные зубы почти никогда не возникают в этих популяциях.
Рисунок 9.14. (A) Если смотреть сверху, хорошо изношенный зубной ряд англосаксонского черепа возрастом 1200 лет показывает очень серьезный износ всех зубов.Это привело к обнажению пульпы первого коренного зуба (стрелки), в результате чего образовался болезненный абсцесс. (B) Вид сбоку того же черепа, что и на (A), показывает место зубного абсцесса (стрелка) под корнями первого коренного зуба. Обратите внимание на сильное уплощение верхних поверхностей коронок.
Источник : © Мелани Николс. Предоставлено Хантерианским музеем Королевского колледжа хирурговРисунок 9.15. Изношенный зубной ряд показывает потерю зубного вещества между соседними зубами, что приводит к очень широкой точке контакта (стрелки).Сравните это с узкой точкой контакта на рисунке 9.12. По мере того, как зубы продвигаются вперед, чтобы закрыть потенциальное пространство (мезиальный дрейф), в задней части челюсти предоставляется достаточно места, чтобы последний коренной зуб прорезывал нормально и не подвергался ударам.
Мягкая, обработанная и приготовленная пища, которую ест большинство современных жителей Северной Америки и Европы, означает, что население не слишком активно пережевывает пищу. Зубы изнашиваются не так сильно и, следовательно, не сильно продвигаются вперед.Это приводит к нехватке места в задней части челюсти для прорезывания зубов мудрости, которые часто подвергаются ретенции (рис. 9.13), что обеспечивает хирургу-стоматологу бесконечный приток пациентов. Возможно, если бы мы все жевали кожу тюленя по полчаса каждый день, ретинированные зубы мудрости остались бы в прошлом!
У людей (и млекопитающих) постоянный набор должен сохраняться, по крайней мере, до достижения репродуктивного возраста, а в идеале — в течение некоторого времени после него. Благодаря достижениям медицины сегодня люди могут легко дожить до 80 лет и более.Особенности, которые эволюционировали, чтобы продлить срок службы зубов млекопитающих, включают их большой размер (особенно у некоторых крупных травоядных, таких как лошадь и корова; см. Главу 4) и наличие очень толстого слоя эмали, самого твердого и стойкого. вещество в природе. У людей слой эмали может иметь максимальную толщину 2,5 мм, тогда как эквивалентный слой, покрывающий дентин у позвоночных, не являющихся млекопитающими, обычно очень тонкий (около 0,2 мм) и имеет более простую структуру. Наличие корней, прикрепленных к костной лунке фиброзной мембраной, позволяет легкими движениями поддерживать контакт с зубом после того, как вещество зуба стерлось.
Нумерация зубов — Bliss Dental Arts San Diego
Представьте себя в кабинете стоматолога с открытым ртом и, вероятно, с нелепым и неудобным выражением лица. Ваш рот испытывает странное ощущение сухости из-за того, что он открыт в течение длительного периода времени, а также неконтролируемое слюноотделение в боковых сторонах рта. У вас болит челюсть, и вам постоянно кажется, что вы падаете на стул. Добавьте к этому, что вы чувствуете себя потерянным, когда пытаетесь понять, как дантист изливает длинный список цифр ассистенту стоматолога, когда протыкают каждый зуб.Этот мучительный пейзаж является фоном для очень важного вопроса, который Меган Хилл задает в своей статье «Зубастая правда»: что эти цифры означают о здоровье ваших зубов?
Нумерация зубов
Когда дантист нумерует ваши зубы, первая цифра представляет собой код, идентифицирующий тип исследуемого зуба (например, резец, клык, моляр и т. Д.). Второе число — это ваш результат в пародонтальной таблице, которая проверяет пародонтоз десен по шкале от 1 до 6.Чем выше число в таблице, тем больше вероятность того, что у вас плохое состояние десен, что может привести к потере зубов. Ваша оценка в таблице определяется стоматологом с помощью инструмента, который протыкается между десной и зубом и оценивает состояние здоровья. Каждое число на самом деле является мерой в миллиметрах, а зуб можно измерить до шести раз. Оценка 1-3 означает, что зуб в целом нормальный и здоровый. Но получение 4 или более баллов означает, что между десной и зубом имеется больший, чем обычно, карман. Более высокий балл также может означать воспаление / кровотечение в деснах и может быть сигналом о том, что вокруг зуба развивается потеря костной массы или пародонтоз.
Заболевания пародонта
По оценкам Британского общества пародонтологии, половина британского населения страдает пародонтозом (также известным как заболевание десен), а 10% на самом деле страдают от наиболее тяжелой формы пародонтоза. Всемирная организация здравоохранения ставит пародонтоз на шестое место среди наиболее распространенных болезней человечества. Установлено, что основными причинами заболевания являются плохая гигиена полости рта, диабет, генетика, употребление табака, плохое питание и стресс.Меган Хилл заканчивает свою статью мудрым советом: «Закажите это обследование!»
Как это:
Нравится Загрузка …
СвязанныеСигнализация мезенхимального Wnt / β-катенина ограничивает количество зубов | Развитие
Каноническая передача сигналов Wnt / β-катенина играет важную роль во время инициации и морфогенеза зубов, а также в дифференцировке зубных клеток (Liu et al., 2008; Lan et al., 2014; Балик и Теслефф, 2015). Wnts представляют собой паракринные сигнальные молекулы ближнего действия, которые регулируют большинство процессов развития, а также обновление стволовых клеток и тканей (Clevers et al., 2014; Farin et al., 2016). Во время раннего морфогенеза зубов лиганды Wnt экспрессируются в зубном эпителии и сигнализируют преимущественно внутри эпителия (Sarkar and Sharpe, 1999; Liu et al., 2008). Передача сигналов Wnt / β-катенина особенно интенсивна в эпителиальных центрах передачи сигналов, включая ранние центры передачи сигналов, называемые узлами инициации (IKs) (Ahtiainen et al., 2016) и эмалевые узлы (EKs), которые регулируют морфогенез почкования и коронки, соответственно (Liu et al., 2008; Ahtiainen et al., 2016; Balic and Thesleff, 2015). Существует также активная передача сигналов β-катенина в мезенхиме, лежащей в основе раннего зубного эпителия, что необходимо для нормального морфогенеза эпителия (Liu et al., 2008; Fujimori et al., 2010; Balic and Thesleff, 2015) и для индукции одонтогенного судьба (Chen et al., 2009). Однако было показано, что повышенные уровни передачи сигналов Wnt / β-катенина в дентальной мезенхиме подавляют одонтогенную судьбу (Liu et al., 2013). Принудительная активация передачи сигналов Wnt в оральном эпителии приводит к непрерывному зарождению новых зубов как у эмбриональных, так и у взрослых мышей (Järvinen et al., 2006; Liu et al., 2008; Wang et al., 2009), тогда как мыши со сверхэкспрессией ингибитора Wnt Dkk1 в зубном эпителии обнаруживает остановку развития зубов на стадии инициации (Andl et al., 2002). В совокупности эти данные убедительно указывают на то, что Wnts являются наиболее передовыми сигналами индукции зубов.
Стоматологические аномалии, в частности те, которые влияют на количество зубов, являются одними из наиболее распространенных отклонений в развитии у людей.Преимущественно они представлены в виде уменьшенного числа зубов, что известно как агенезия зубов или гиподонтия. Во многих случаях генные мутации, вызывающие фенотип, были раскрыты (Arte et al., 2013; Mues et al., 2014; Yin and Bian, 2015), но лежащие в основе молекулярные механизмы часто остаются нерешенными. Тяжелая агенезия зубов, или олигодонтия, часто связана с различными человеческими синдромами, чаще всего с дефектами других эктодермальных органов (Lefebvre and Mikkola, 2014; Plaisancié et al., 2013).Агенез зубов часто вызывается мутациями в компонентах сигнального пути Wnt / β-catenin, наиболее распространенным геном является WNT10A (van den Boogaard et al., 2012; Arte et al., 2013; Plaisancié et al., 2013). Интересно, что гетерозиготные мутации с потерей функции в ингибиторе обратной связи Wnt AXIN2 (Lustig et al., 2002; Jho et al., 2002) также приводят к тяжелому агенезу зубов (Lammi et al., 2004; Marvin et al. , 2011; Wong et al., 2014; Yue et al., 2016; OMIM 608615), что указывает на негативную роль передачи сигналов Wnt в формировании зубов.Человеческие AXIN2 мутации особенно ингибируют образование сменных зубов и задних коренных зубов, но не оказывают значительного влияния на развитие молочных (первичных) зубных рядов (Lammi et al., 2004). Фенотип гиподонтии AXIN2 подразумевает, что усиление передачи сигналов Wnt / β-catenin может также подавлять формирование зубов. Во время развития зубов Axin2 экспрессируется в эпителиальных EKs и в дентальной мезенхиме (Lammi et al., 2004). Поскольку генетическая активация передачи сигналов Wnt / β-катенина в оральном эпителии мышей (Järvinen et al., 2006; Лю и др., 2008; Wang et al., 2009) индуцирует огромное развитие дополнительных зубов, нарушение формирования зубов у людей, несущих мутации AXIN2 , скорее всего, вызвано усилением передачи сигналов Wnt / β-catenin в мезенхиме.
Противоположный стоматологический фенотип наблюдается при клейдокраниальной дисплазии, синдроме человека, при котором гетерозиготные мутации потери функции в транскрипционном факторе RUNX2 вызывают образование нескольких сверхкомплектных зубов или гипердонтию.Дополнительные зубы развиваются как дополнительные замещающие зубы и задние моляры (Jensen and Kreiborg, 1990; Lee et al., 1997; Jaruga et al., 2016; OMIM 119600). Во время развития зубов функция Runx2 регулируется с помощью Fgfs (Åberg et al., 2004), но исследования развивающихся костей показали, что передача сигналов Wnt / β-catenin положительно регулирует Runx2 (Komori, 2011).
Постоянный зубной ряд человека формируется посредством процесса, называемого последовательным формированием зубов, при котором новый зуб формируется из продолжения зубной пластинки предшествующего зуба (Juuri et al., 2013; Джури и Балич, 2017). Есть два режима последовательного формирования зубов, которые происходят во время формирования постоянного зубного ряда человека: замена зубов, при которой образуются вторичные зубы; и последовательное добавление зубов, так формируются задние моляры. Мыши не заменяют свои зубы, но мы ранее показали, что формирование их задних коренных моляров (M2 и M3) имеет общие морфологические, а также молекулярные характеристики с инициированием замены зубов у эмбрионов человека и хорька.В частности, они развиваются из Sox2-экспрессирующих предшественников в дентальной пластинке (Järvinen et al., 2009; Jussila et al., 2014; Juuri et al., 2013). Таким образом, мыши M2 и M3 могут быть использованы в качестве модели для изучения последовательного формирования зубов и молекулярных механизмов стоматологических аномалий, влияющих на постоянный зубной ряд при синдромах человека.
В настоящей работе мы исследовали эффекты модуляции сигнала Wnt / β-катенина на последовательное формирование моляров на моделях мышей и в культурах ex vivo .Мы увеличили экспрессию β-катенина in vivo, , экспрессируя стабилизированную форму β-катенина, и in vitro, , добавив BIO, ингибитор GSK, к культивируемым зубам in vitro. Помимо того, что он является внутриклеточным эффектором передачи сигналов Wnt, β-катенин также участвует в клеточной адгезии, образуя комплексы с кадгеринами на клеточных мембранах, преимущественно в эпителиальных тканях. Однако консенсус в области β-catenin, по-видимому, заключается в том, что экспериментальное увеличение цитоплазматического β-catenin имитирует эффект усиленной передачи сигналов Wnt / β-catenin (Fagotto, 2013; McCrea and Gottardi, 2016).Мы предполагаем, что эффекты повышенного β-catenin в зубной мезенхиме являются результатом усиления передачи сигналов Wnt, а не увеличения адгезии клеток. Это подтверждается нашим открытием, что нет очевидных изменений в плотности мезенхимальных клеток в гистологических срезах первых моляров, культивируемых с BIO.
Мы демонстрируем, что уровень мезенхимальной передачи сигналов Wnt / β-катенина имеет решающее значение для инициирования последовательного формирования зубов: увеличение активности Wnt / β-катенина в дентальной мезенхиме подавляло развитие задних моляров, тогда как снижение мезенхимального Wnt / β-катенина был связан с их непрерывным развитием.Наши результаты также предполагают, что у людей врожденное отсутствие зубов и образование лишних зубов, вызванное мутациями в AXIN2 и RUNX2 , соответственно, являются результатом модуляции передачи сигналов Wnt / β-catenin в зубной мезенхиме.
Ранее мы показали, что стабилизация β-катенина в оральном эпителии эмбрионов мышей (β-cat Δex3K14 / + ) приводит к непрерывному зарождению новых зубов (Järvinen et al., 2006). Здесь мы использовали эту мышиную модель, чтобы исследовать, связано ли последовательное формирование зубов с уровнем передачи сигналов мезенхимы Wnt. Иммунофлуоресцентный анализ фосфорилированного β-катенина в контрольных молярах E16 показал интенсивное окрашивание β-катенина, локализованное на поверхности клеток в большинстве эпителиальных клеток (рис. 1A). Окрашивание ядер наблюдалось во внутреннем эпителии эмали, особенно в местах вторичных ЭК (рис. 1А) и в зубной мезенхиме, лежащей под зубным эпителием (звездочка на рис.1А). Анализ срезов моляров β-cat Δex3K14 / + на E16 показал несколько очагов интенсивного окрашивания ядерного β-катенина (рис. 1B), что согласуется с нашей более ранней демонстрацией экспрессии BAT-gal в этих мутанты (Järvinen et al., 2006). Ранее мы сообщали, что эти фокусы экспрессируют несколько маркеров ЭК, включая Shh , Fgf4 , Wnt10a и Edar (Järvinen et al., 2006; Fig. 1C, D). Эти индуцированные эпителиальные сигнальные центры, вероятно, представляют собой IK, недавно описанные в зубных плакодах во время зарождения зуба (Ahtiainen et al., 2016). Интересно, что окрашивание β-катенина полностью отсутствовало в мезенхимальных клетках моляров β-cat Δex3K14 / + (звездочка на рис. 1B).
Рис. 1.
Передача сигналов мезенхимального Wnt / β-катенина отсутствует во время непрерывного образования зубцов у мутантов β-cat Δex3K14 / + . (A, B) Иммуноокрашивание β-катенина. (C-N) In situ гибридизационный анализ экспрессии Shh и ингибиторов Wnt / β-катенина в контроле и E16 β-cat Δex3K14 / + челюстей.(В, Д) Шш . (E-H) Ингибиторы обратной связи (репортеры Wnt): Axin2, и Drapc1. (I-N) Ингибиторы: Sostdc1 , Dkk1 и Dkk4. Масштабные линейки: 100 мкм. д — эпителий; ЭК, эмалевый узел; белая звездочка указывает на мезенхиму; пунктирная линия отделяет зубной эпителий от окружающей мезенхимы.
Рис. 1.
Передача сигналов мезенхимального Wnt / β-катенина отсутствует во время непрерывного образования зубцов у мутантов β-cat Δex3K14 / + . (A, B) Иммуноокрашивание β-катенина. (C-N) In situ гибридизационный анализ экспрессии Shh и ингибиторов Wnt / β-катенина в контроле и E16 β-cat Δex3K14 / + челюстей. (В, Д) Шш . (E-H) Ингибиторы обратной связи (репортеры Wnt): Axin2, и Drapc1. (I-N) Ингибиторы: Sostdc1 , Dkk1 и Dkk4. Масштабные линейки: 100 мкм. д — эпителий; ЭК, эмалевый узел; белая звездочка указывает на мезенхиму; пунктирная линия отделяет зубной эпителий от окружающей мезенхимы.
Axin2 и Drapc1 , мишени и ингибиторы обратной связи передачи сигналов Wnt / β-катенина (Behrens et al., 1998; Shimomura et al., 2010), были экспрессированы в мезенхиме сосочков зубов, ближайшей к эпителию эмали в контрольных зубах. на E16, как продемонстрировано анализом гибридизации in situ (рис. 1E, G). Экспрессия этих генов полностью отсутствовала в зубной мезенхиме зубных зачатков E16 β-cat Δex3K14 (рис.1F, H), тогда как в эпителии они демонстрировали интенсивные и ограниченные домены экспрессии, соответствующие индуцированным эпителиальным центрам передачи сигналов, то есть IKs.
In situ анализ гибридизации ингибиторов Wnt Sostdc1 и Dkk1 продемонстрировал их экспрессию в зубном сосочке контрольных зубных зачатков E16, за исключением его коронарной (верхней) части, непосредственно подстилающей внутренний эпителий эмали, при этом также экспрессируется Sostdc1 в мезенхиме зубного фолликула, окружающего эпителиальный эмалевый орган (рис.1И, К). В мутантах β-cat Δex3K14 / + мезенхимальные домены экспрессии этих генов простирались до зубного эпителия, при этом Sostdc1 также присутствует в недентальной мезенхиме, подчеркивающей эпителий полости рта (рис. 1J, L). .
Другой ингибитор Wnt, Dkk4 , экспрессировался в EK в зубном эпителии контрольных зубов (рис. 1M), а сильная экспрессия наблюдалась в индуцированных IK у мутантов β-cat Δex3K14 / + (рис.1N). Домены экспрессии β-катенина и проанализированные гены показали, что передача сигналов Wnt / β-катенина ингибируется в мезенхиме β-cat Δex3K14 / + и что это ингибирование происходит ниже принудительной передачи сигналов β-катенина в эпителии. Следовательно, подавление мезенхимальной передачи сигналов Wnt может быть важным для непрерывного образования зубов у мутантов β-cat Δex3K14 / + .
Затем мы использовали систему культивирования органов ex vivo , чтобы проверить, может ли усиленная мезенхимальная передача сигналов Wnt / β-катенин ингибировать непрерывное образование зубов у мутантных мышей β-cat Δex3K14 / + .Мы активировали передачу сигналов Wnt / β-катенина в рассеченных зачатках зубов путем добавления ингибитора GSK BIO в культуральную среду, тем самым предотвращая деградацию β-катенина. Зачатки коренных зубов вырезали из мутантных эмбрионов E13.5 β-cat Δex3K14 / + и культивировали в течение 6 дней с или без BIO и ежедневно фотографировали. В качестве контроля зачатки зубов E13.5 от контрольных однопометников культивировали в течение 6 дней без BIO (рис. 2A, D, G).
Рис.2.
Стимуляция передачи сигналов Wnt / β-катенина с помощью BIO предотвращает непрерывное образование зубов у β-cat Δex3K14 / + мутантов ex vivo . (A) Фотография контрольного зубного зачатка E13.5, культивированного в течение 6 дней. М1 и М2 развиваются. (B) Множественные маленькие зубы, сформированные из одного зубного зачатка мутантного эмбриона E13.5 β-cat Δex3K14 / + после 6 дней культивирования в контрольной среде ( n = 9). (C) BIO (2,0 мкМ) предотвращал образование новых зубов в мутантном эксплантате β-cat Δex3K14 / + ( n = 11).(D-F) Гистологические срезы эксплантатов (окрашивание гематоксилином и эозином). (G-I) Иммунофлуоресцентная локализация β-катенина. (G) β-Катенин преимущественно локализуется в эпителии, тогда как в мезенхиме не наблюдается окрашивания. (H) Интенсивное очаговое окрашивание β-катенина обнаруживается в эпителии необработанного эксплантата β-cat Δex3K14 / + , но окрашивание мезенхимы отсутствует. (I) Обработка BIO приводит к усилению окрашивания β-катенина в мезенхиме зубов. Масштабные линейки: 100 мкм. Белая звездочка указывает на мезенхиму.
Рис. 2.
Стимуляция передачи сигналов Wnt / β-catenin с помощью BIO предотвращает непрерывное образование зубов у β-cat Δex3K14 / + мутантов ex vivo . (A) Фотография контрольного зубного зачатка E13.5, культивированного в течение 6 дней. М1 и М2 развиваются. (B) Множественные маленькие зубы, сформированные из одного зубного зачатка мутантного эмбриона E13.5 β-cat Δex3K14 / + после 6 дней культивирования в контрольной среде ( n = 9). (C) BIO (2,0 мкМ) предотвращал образование новых зубов в мутантном эксплантате β-cat Δex3K14 / + ( n = 11).(D-F) Гистологические срезы эксплантатов (окрашивание гематоксилином и эозином). (G-I) Иммунофлуоресцентная локализация β-катенина. (G) β-Катенин преимущественно локализуется в эпителии, тогда как в мезенхиме не наблюдается окрашивания. (H) Интенсивное очаговое окрашивание β-катенина обнаруживается в эпителии необработанного эксплантата β-cat Δex3K14 / + , но окрашивание мезенхимы отсутствует. (I) Обработка BIO приводит к усилению окрашивания β-катенина в мезенхиме зубов. Масштабные линейки: 100 мкм. Белая звездочка указывает на мезенхиму.
Как показано ранее (Järvinen et al., 2006), новые зубы непрерывно зарождались в эксплантах β-cat Δex3K14 / + во время культивирования в контрольной среде (Рис. 2B, E). В присутствии BIO эксплантаты β-cat Δex3K14 / + демонстрировали ингибирование последовательного зачатия зуба дозозависимым образом, где более низкая доза (2,0 мкМ) предотвращала зачатие зуба в 5/12 эксплантатах, а более высокая доза (10 мкМ) предотвращает зарождение зубов во всех четырех эксплантах (4/4) (рис.2C, F и данные не показаны). Активация передачи сигналов Wnt / β-катенина была подтверждена иммуноокрашиванием ядерного β-катенина в мезенхимальных клетках в эксплантатах, культивируемых с BIO (3,75 мкМ) (рис. 2I). Эксплантаты β-cat Δex3K14 / + , культивированные в контрольной среде, продемонстрировали интенсивное очаговое иммуноокрашивание ядерного β-катенина в эпителиальных центрах передачи сигналов формирующихся дополнительных зубов, тогда как окрашивание β-катенина полностью отсутствовало в мезенхиме (рис. 2H). ). Вместе эти данные указывают на то, что усиление передачи сигналов мезенхимального Wnt / β-catenin может ингибировать инициацию зуба.
Далее мы хотели проанализировать эффект повышенной передачи сигналов мезенхимального Wnt / β-catenin на последовательное формирование зубов in vivo . Развитие задних моляров M2 и M3 у мышей представляет собой процесс последовательного формирования зубов и напоминает замену зубов как морфологически, так и молекулярно (Juuri et al., 2013). Мы получили мышей, у которых экспрессия стабилизированного β-катенина была направлена на эмбриональную мезенхиму с использованием промотора Dermo1Cre .Эмбрионы βcat Δex3Dermo / + , однако, умерли пренатально до того, как M1 достиг стадии зачатка и до начала развития M2. Поэтому мы препарировали развивающиеся зубные зачатки из мутантных и контрольных эмбрионов на E12.5 и культивировали их ex vivo для анализа морфогенеза M1 и образования M2. Хотя и контрольные, и мутантные зубные зачатки M1 росли в культуре, ни один из них не развился дальше ранней стадии колокола, и M2 не смог образоваться в обоих (не показано). Скорее всего, это было связано с ранней стадией развития на заре культуры.
Затем мы трансплантировали рассеченный контроль E12.5 и мутантные зубные зачатки M1 βcat Δex3Dermo / + под капсулу почки взрослых голых мышей и собирали ткани через 3 недели. Количество сформированных зубов было разным. Контрольные эксплантаты ( n = 9) образовывали от одного до трех моляров следующим образом: в одном экспланте — все три моляра; в трех эксплантах — два моляра; в трех эксплантах — один коренной зуб; а в двух эксплантатах — без зубов.Мутантные эксплантаты βcat Δex3Dermo / + ( n = 11) дали один моляр: у семи эксплантов образовался только один моляр; у четырех эксплантатов зубы не образовывались (рис. 3). Следовательно, способность генерировать более одного моляра наблюдалась только в контроле, что позволяет предположить, что активация передачи сигналов Wnt / β-catenin в дентальной мезенхиме приводит к ингибированию последовательного образования зубов.
Рис.3.
Принудительная активация передачи сигналов Wnt / β-катенина в эмбриональной мезенхиме (βcat Δex3Dermo / + ) ингибирует образование M2 и M3. (A-F) Зубы, полученные из контроля и βcat Δex3Dermo / + мутантных зачатков коренных зубов. Зачатки коренных зубов вырезали из челюстей эмбриона E12.5 и выращивали под капсулой почки голых мышей в течение 3 недель. (A-C) Изображения коренных зубов, полученных из зачатков контрольных зубов. Развиваются от одного до трех моляров ( n = 9). (D-F) Изображения зубов, полученных из мутантных эксплантатов.У мутантов развился только M1 ( n = 11). Масштабные линейки: 1 мм.
Рис. 3.
Принудительная активация передачи сигналов Wnt / β-катенина в эмбриональной мезенхиме (βcat Δex3Dermo / + ) ингибирует образование M2 и M3. (A-F) Зубы, полученные из контроля и βcat Δex3Dermo / + мутантных зачатков коренных зубов. Зачатки коренных зубов вырезали из челюстей эмбриона E12.5 и выращивали под капсулой почки голых мышей в течение 3 недель. (A-C) Изображения коренных зубов, полученных из зачатков контрольных зубов.Развиваются от одного до трех моляров ( n = 9). (D-F) Изображения зубов, полученных из мутантных эксплантатов. У мутантов развился только M1 ( n = 11). Масштабные линейки: 1 мм.
Эмбриональная летальность βcat Δex3Dermo / + мышей препятствует дальнейшему анализу принудительной передачи сигналов Wnt / β-catenin в мезенхиме и его роли в последовательном формировании моляров in vivo .Поэтому мы использовали ex vivo культур органов, чтобы изучить влияние усиленной передачи сигналов Wnt / β-катенин на образование M2 у контрольных мышей. M2 развивается из заднего продолжения эпителия M1, также называемого задней зубной пластиной, задней частью хвоста (кончик) и непрерывной пластиной (Juuri et al., 2013; Gaete et al., 2015; Juuri and Balic, 2017). Мы вскрыли зародыши зуба M1 эмбриона мыши на стадии, когда M2 инициирован (E14.5), и непрерывная пластинка, которая указывает на начальную точку развития M2, четко видна (рис.4А). Эксплантаты культивировали и наблюдали за их развитием в присутствии или в отсутствие BIO в течение 6 дней.
Рис. 4.
Стимуляция передачи сигналов β-катенина ex vivo ингибирует образование M2. (A, C) E14,5 моляров (M1) из эмбрионов мыши TCF / Lef: h3B-GFP в начале культивирования. Звездочки указывают на непрерывную зубную пластинку (хвост) M1, которая дает начало M2.(B) M2, образованный в контрольной среде после 6 дней культивирования (стрелка) ( n = 32). (D) BIO (2,0 мкМ) ингибирует образование M2 (стрелка), тогда как M1 меньше с более мелкими выступами ( n = 29). Масштабные линейки: 100 мкм.
Рис. 4.
Стимуляция передачи сигналов β-катенина ex vivo ингибирует образование M2. (A, C) E14,5 моляров (M1) из эмбрионов мыши TCF / Lef: h3B-GFP в начале культивирования. Звездочки указывают на непрерывную зубную пластинку (хвост) M1, которая дает начало M2.(B) M2, образованный в контрольной среде после 6 дней культивирования (стрелка) ( n = 32). (D) BIO (2,0 мкМ) ингибирует образование M2 (стрелка), тогда как M1 меньше с более мелкими выступами ( n = 29). Масштабные линейки: 100 мкм.
Мы использовали Lef1 -репортерных мышей для облегчения наблюдения за развитием моляров во время культивирования. Морфогенез коронки зуба связан с появлением Lef1 -выражающих очагов, т.е.е. первичные и вторичные ЭК, обозначающие начало коронки и формирование бугорка зуба соответственно (рис. 4А, В). В контрольной среде M2 образуется к 6-му дню у большинства эксплантов (32/40) (желтая стрелка на фиг. 4B). Добавление BIO ингибировало образование M2 дозозависимым образом. При высоких концентрациях BIO (3,75 мкМ-15,0 мкМ) развитие M2 подавлялось во всех эксплантах (31/31), а при низкой концентрации (2,0 мкМ) в большинстве эксплантов (29/37) (желтая стрелка на рис. 4D) . M1 демонстрировал аномальный морфогенез коронки при высоких концентрациях BIO и при низкой концентрации (2.0 мкМ) коронки были меньше, чем в контроле, а бугорки неглубокие (рис. 4D). Следовательно, 2,0 мкМ BIO использовали в следующих экспериментах.
Затем мы исследовали нижестоящие эффекторы и молекулярные механизмы, связанные с задержкой инициации зуба, вызванной усилением передачи сигналов Wnt / β-catenin. Axin2 является мишенью и хорошо зарекомендовавшим себя ингибитором с обратной связью пути Wnt / β-катенин, а мутации AXIN2 человека ингибируют образование замещающих зубов и задних моляров, которые образуются в результате последовательного формирования зубов.Поэтому мы решили сначала изучить роль Axin2 в инициации M2 и его возможный вклад в наблюдаемые нами фенотипы. Мы культивировали зубные зародыши E14.5 M1 дикого типа в течение 2 дней, разделяли M1 и M2 и проводили количественную ПЦР в реальном времени (RT-qPCR) на контрольном и обработанном BIO M2. Ингибирование развития M2 с помощью BIO было связано с двукратным увеличением экспрессии Axin2 (фиг. 5A).
Рис.5. Экспрессия
Axin2 и Drapc1 повышена в мутантной дентальной мезенхиме Runx2 . (A) Разделение E14.5 M1 и M2 после 2 дней культивирования для анализа RT-qPCR. Стимуляция экспрессии Axin2 с помощью BIO (2 мкМ) в M2 ( t -тест). (B) Выражение Axin2 , Lef1, и Runx2 в сагиттальных сечениях на E14.5 и E15.5 в M1 и M2. Узлы эпителиальной эмали экспрессируют Axin2 , но в остальном экспрессия в значительной степени ограничивается мезенхимой.На E14.5 экспрессия Axin 2 лежит в основе хвоста эпителия M1 в том месте, где инициируется M2 (белая звездочка). На E15.5 мезенхимальный зубной сосочек, мезенхима, окружающая M1 и M2, и мезенхима, лежащая в основе орального эпителия, экспрессируют Axin2 . (C) Локализация Axin2 и Drapc1 в мезенхиме мутантных зубов Runx2 -null и контрольных зубов из однопометников на E13.5. Пунктирная линия (B, C) отделяет зубной эпителий от окружающей мезенхимы.(D) Экспрессия Axin2, и Drapc1 активируется в мезенхиме нулевого мутанта Runx2- и гетерозиготных зубных зачатков Runx2 на E13.5 ( t -тест). * P <0,05, *** P <0,0005. Масштабные линейки: 100 мкм.
Рис. 5.
Axin2 и Экспрессия Drapc1 повышена в мутантной дентальной мезенхиме Runx2 . (A) Разделение E14.5 M1 и M2 после 2 дней культивирования для анализа RT-qPCR.Стимуляция экспрессии Axin2 с помощью BIO (2 мкМ) в M2 ( t -тест). (B) Выражение Axin2 , Lef1, и Runx2 в сагиттальных сечениях на E14.5 и E15.5 в M1 и M2. Узлы эпителиальной эмали экспрессируют Axin2 , но в остальном экспрессия в значительной степени ограничивается мезенхимой. На E14.5 экспрессия Axin 2 лежит в основе хвоста эпителия M1 в том месте, где инициируется M2 (белая звездочка). На E15.5 мезенхимальный зубной сосочек, мезенхима, окружающая M1 и M2, и мезенхима, лежащая в основе орального эпителия, экспрессируют Axin2 .(C) Локализация Axin2 и Drapc1 в мезенхиме мутантных зубов Runx2 -null и контрольных зубов из однопометников на E13.5. Пунктирная линия (B, C) отделяет зубной эпителий от окружающей мезенхимы. (D) Экспрессия Axin2, и Drapc1 активируется в мезенхиме нулевого мутанта Runx2- и гетерозиготных зубных зачатков Runx2 на E13.5 ( t -тест). * P <0,05, *** P <0.0005. Масштаб: 100 мкм.
Как сообщалось ранее (Lammi et al., 2004; Lohi et al., 2010), Axin2 экспрессируется в дентальной мезенхиме и EKs во время морфогенеза зубов (Fig. 5B). В сагиттальных срезах челюстей во время инициации M2 (E14.5) Axin2 экспрессия была интенсивной в дентальной мезенхиме, лежащей в основе эпителия (Fig. 5B). Lef1 , хорошо известный маркер передачи сигналов Wnt / β-катенин, коэкспрессировался с Axin2 в дентальной мезенхиме (рис.5Б). На E14.5 экспрессия Axin2 была обнаружена также в мезенхиме, лежащей в основе заднего хвоста M1 (континуальная пластинка), т.е. в том месте, где инициируется M2 (фиг. 5B, звездочка). На E15.5 интенсивная экспрессия Axin2 присутствовала в M1 и M2 в мезенхиме зубного сосочка, лежащей под зубным эпителием, а в M2 сильная мезенхимальная экспрессия окружала весь зубной эпителий и продолжалась сзади под эпителием полости рта вплоть до верхних моляров (рис. .5Б). Эти результаты показывают, что существует мезенхимальная сигнальная активность Wnt / β-catenin вокруг развивающегося эпителия M2, которая, скорее всего, усиливается BIO в наших экспериментах.
Мыши с нулевым мутантом Axin2 проявляют повышенную чувствительность к передаче сигналов Wnt / β-catenin и широко используются для анализа пути Wnt. У этих мышей наблюдается довольно легкий дефект развития костей черепа (Yu et al., 2005). Мы проанализировали зубные ряды взрослых нулевых мутантных мышей Axin2- ( n = 10) под стереомикроскопом и не обнаружили каких-либо отклонений по сравнению с контрольными однопометниками. Все зубы присутствовали, их размер, форма, а также структура эмали были нормальными (рис. S1). Эти находки показывают, что у мышей (в отличие от людей) делеция функции Axin2 не оказывает значительного влияния на развитие зубов.
Мы исследовали, повлияла ли потеря Axin2 на экспрессию выбранных регуляторных генов зубов с помощью гибридизационного анализа in situ в эмбриональных Axin2- нулевых мутантных зубах. Shh , Fgf4 , Lef1 , Fgf3 , Dkk1 , Runx2 и Drapc1 были выражены в нормальных формах и интенсивности в молярах E13.5 и E14.5 (не показаны). Интересно, что Axin2 -null дентальные ткани были более чувствительны к стимуляции передачи сигналов Wnt / β-catenin во время дифференцировки дентина и эмаль-образующих клеток. Когда нулевые мутантные эксплантаты зубов E13.5 Axin2 культивировали в присутствии BIO (10 мкМ), дифференцировка эпителиальных клеток в амелобласты, а также мезенхимальных клеток в одонтобласты ускорялась (рис.S2). Отсутствие морфологических изменений у мутантных мышей Axin2 указывает на то, что функция Axin2 может быть компенсирована другими антагонистами Wnt или ингибиторами обратной связи, включая Drapc1.
Фенотип, вызванный мутациями AXIN2 в зубных рядах человека, т.е. тяжелая агенезия зубов (oligodontia), влияет преимущественно на последовательно формирующиеся зубы (Lammi et al., 2004) и противоположен фенотипу, возникающему в результате мутаций RUNX2 , вызывающих последовательное развитие множественных дополнительных зубов при синдроме кледокраниальной дисплазии (Jensen and Kreiborg, 1990; Lee et al., 1997). Интересно, что Axin2 был указан как прямой ген-мишень для Runx2 в кости (Li et al., 2009; McGee-Lawrence et al., 2013). Мы сравнили паттерны экспрессии Axin2 и Runx2 (фиг. 5B) во время образования M2 и исследовали экспрессию Axin2 у нулевых мутантных мышей Runx2-. Runx2 обнаруживает очевидную совместную локализацию с Axin2 в мезенхиме, окружающей M1 и M2 на E15.5 (Fig. 5B). Поскольку молярное развитие (M1) приостанавливается у мутантных эмбрионов Runx2 между E13 и E14 (до перехода от стадии зачатка к стадии шляпки) (Åberg et al., 2004), мы исследовали экспрессию Axin2 в нуль-мутантах Runx2- на E13.5. In situ гибридизационный анализ показал немного более высокую экспрессию Axin2 в нулевых мутантных зубах Runx2- по сравнению с контрольными зубами однопометников (фиг. 5C). Анализ RT-qPCR экспрессии Axin2 в дентальной мезенхиме E13.5 Runx2-null показал, что Axin2 экспрессировался на значительно более высоких уровнях ( P = 0,0005) в мутантной дентальной мезенхиме Runx2 по сравнению с контрольными однопометниками. (Инжир.5D).
Подобно Axin2 , Drapc1 является ингибитором обратной связи передачи сигналов β-катенина, и его характер экспрессии был подобен Axin2 в контроле и β-cat Δex3K14 / + зубных зачатков (рис. 1). Таким образом, мы исследовали, регулируется ли экспрессия Drapc1 с помощью Runx2. Действительно, как гибридизация in situ , так и анализ RT-qPCR показали повышенную экспрессию Drapc1 в мезенхиме нулевых мутантных зубных зачатков Runx2- по сравнению с контролем ( P = 0.035) (Рис. 5C, D).
Чтобы изучить функцию Runx2 во время последовательного формирования зубцов, мы проанализировали сигнальную активность Wnt / β-катенина во время инициации M2 у гетерозигот Runx2 , поскольку остановка морфогенеза M1 на стадии зачатка отключает этот анализ в Runx2- нулевые эмбрионы. Анализ qPCR продемонстрировал значительное увеличение экспрессии Axin2 и Drapc1 в мезенхиме гетерозиготных зубов Runx2 на ст. E14.5, что свидетельствует об увеличении активности Wnt / β-катенина. Затем мы локализовали сигнальную активность Wnt / β-catenin, используя репортер Lef1-GFP и ядерное окрашивание β-catenin, в гистологических срезах на E14.5 у гетерозигот Runx2 и контрольных однопометников (рис. 6). В контрольных зубах клетки с активной передачей сигналов Wnt были обнаружены в мезенхиме, смежной с EK (Fig. 6C-F), и только несколько клеток с активной передачей сигналов Wnt были обнаружены под континуальной пластинкой (Fig. 6K-M). Напротив, в гетерозиготных зубах количество клеток с активной передачей сигналов Wnt было значительно ниже (рис.6G-J), особенно под непрерывной пластинкой, где не было обнаружено клеток с активной передачей сигналов Wnt (Fig. 6N-P).
Рис. 6.
Передача сигналов Wnt / β-catenin ингибируется в Runx2 гетерозиготе в мезенхиме под эмалевым узлом M1 и континуальной пластинкой, формирующей M2. (A, B) Окрашивание Lef1-GFP и ядерного β-катенина указывает на области активной передачи сигналов Wnt в контроле Runx2 (A) и Runx2 +/- (B).(C-F) Более высокое увеличение контрольного зуба с активной передачей сигналов Wnt в мезенхиме под эмалевым узлом (стрелки). (G-J) Runx2 +/- с меньшей передачей сигналов Wnt в мезенхимных клетках под эмалевым узлом (стрелка). (K-M) Под контролем непрерывной пластинки находится несколько клеток с активной передачей сигналов Wnt (стрелки). (N-P) Под Runx2 +/- непрерывная пластинка, нет клеток с активной передачей сигналов Wnt (стрелки). ЭК, эмалевый узел; CL, сплошная пластинка. Масштабные линейки: 100 мкм.
Рис. 6.
Передача сигналов Wnt / β-catenin ингибируется в Runx2 гетерозиготе в мезенхиме под эмалевым узлом M1 и непрерывной пластинкой, формирующей M2. (A, B) Окрашивание Lef1-GFP и ядерного β-катенина указывает на области активной передачи сигналов Wnt в контроле Runx2 (A) и Runx2 +/- (B). (C-F) Более высокое увеличение контрольного зуба с активной передачей сигналов Wnt в мезенхиме под эмалевым узлом (стрелки). (G-J) Runx2 +/- с меньшей передачей сигналов Wnt в мезенхимных клетках под эмалевым узлом (стрелка).(K-M) Под контролем непрерывной пластинки находится несколько клеток с активной передачей сигналов Wnt (стрелки). (N-P) Под Runx2 +/- непрерывная пластинка, нет клеток с активной передачей сигналов Wnt (стрелки). ЭК, эмалевый узел; CL, сплошная пластинка. Масштабные линейки: 100 мкм.
Отсутствие зубного фенотипа и неизменная экспрессия различных генов, связанных с путём Wnt / β-catenin у мышей с нулевым мутантом Axin2 , предполагает, что другие пути передачи сигналов также могут быть задействованы.Мы решили проанализировать экспрессию выбранных генов в BIO-обработанных (2 мкМ) эксплантатах зубов, чтобы точно определить сигнальные пути, связанные с задержкой образования M2. Основываясь на предыдущей литературе по инициированию зуба и нашей недавней работе, показывающей, что усиление передачи сигналов мезенхимального β-катенина опосредуется Fgf10 в нише стволовых клеток зуба, мы сосредоточились на путях Fgf и Bmp (Jussila and Thesleff, 2012; Yang et al., 2015). Мы исследовали BIO-обработанные и контрольные эксплантаты ( Lef1 -репортерные мыши) после 2 дней культивирования на экспрессию потенциальных лигандов и мишеней. Анализ гибридизации in situ контрольных культур E15.5 показал, что Fgf10 и Bmp4 экспрессировались в зубной мезенхиме, тогда как их соответствующие мишени Dusp6 и Id1 экспрессировались в эпителии (рис. 7A, а не показаны), но не было очевидной разницы в интенсивности экспрессии между BIO-обработанными и контрольными эксплантами. Поэтому мы провели RT-qPCR на контрольном и обработанном BIO M2 через 2 дня культивирования (рис.7Б). Повышенные уровни Bmp4 и Id1 , а также подавление экспрессии Fgf10 было обнаружено в BIO-обработанном M2 (фиг. 7B) . Кроме того, были активированы , Fgf2 , Fgf3 и цель Fgf Dusp6 (рис. 7B).
Рис. 7.
Влияние BIO на передачу сигналов Bmp и Fgf и на образование M2. (A) In situ анализ гибридизации экспрессии Fgf10 , Bmp4 , Dusp6 и Id1 в E14.5 моляров через 2 дня культивирования с BIO и без него. Не было обнаружено значительных различий между контрольными культурами и культурами, подвергнутыми воздействию BIO, в характере экспрессии и интенсивности. Масштабные линейки: 100 мкм. (B) После 1 дня культивирования M1 и M2 были разделены для анализа RT-qPCR влияния BIO на экспрессию гена в M2. Bmp4 , Id1 , Fgf2 , Fgf3 и Экспрессия Dusp6 была увеличена в BIO-обработанных эксплантатах M2, тогда как экспрессия Fgf10 была снижена ( t -test).* P <0,05. (C) Моляры E14,5 из Tcf / Lef1 репортерных мышиных эмбрионов культивировали с и без BIO (2,0 мкМ), Bmp4 (100 нг / мл) и Noggin (300 нг / мл) в течение 6 дней для изучения развития М2. Количество образовавшихся M2 / общее количество эксплантов указано на рисунке. BIO и Bmp4 ингибировали образование M2 (красная звездочка), тогда как Noggin не действовал. Белыми звездочками отмечены участки непрерывной зубной пластинки.
Рис. 7.
Влияние BIO на передачу сигналов Bmp и Fgf и на образование M2. (A) In situ анализ гибридизации экспрессии Fgf10 , Bmp4 , Dusp6 и Id1 в молярах E14.5 после 2 дней культивирования с BIO и без него. Не было обнаружено значительных различий между контрольными культурами и культурами, подвергнутыми воздействию BIO, в характере экспрессии и интенсивности. Масштабные линейки: 100 мкм. (B) После 1 дня культивирования M1 и M2 были разделены для анализа RT-qPCR влияния BIO на экспрессию гена в M2. Bmp4 , Id1 , Fgf2 , Fgf3 и Экспрессия Dusp6 была увеличена в BIO-обработанных эксплантатах M2, тогда как экспрессия Fgf10 была снижена ( t -test).* P <0,05. (C) Моляры E14,5 из Tcf / Lef1 репортерных мышиных эмбрионов культивировали с и без BIO (2,0 мкМ), Bmp4 (100 нг / мл) и Noggin (300 нг / мл) в течение 6 дней для изучения развития М2. Количество образовавшихся M2 / общее количество эксплантов указано на рисунке. BIO и Bmp4 ингибировали образование M2 (красная звездочка), тогда как Noggin не действовал. Белыми звездочками отмечены участки непрерывной зубной пластинки.
Эти данные показали, что передача сигналов Bmp может играть агонистическую роль с передачей сигналов Wnt / β-catenin при инициации M2.Для дальнейшего анализа влияния Bmp4 на развитие M2 мы использовали культуры эксплантатов зубных зачатков E14.5 и обработали их рекомбинантным белком Bmp4 (100 нг / мл). M2 образовывался в большинстве эксплантов, выращенных в контрольной среде в течение 6 дней (39/49), но только в 14/40 образцов, обработанных Bmp4 (фиг. 7C). Это было сопоставимо с частотой развития M2 в образцах, обработанных BIO. Ингибитор Bmp Noggin (300 нг / мл) не оказывал значительного влияния на образование M2 (18/21, фиг. 7D). При добавлении вместе с BIO Noggin не спасал образование M2 (не показано).
Передача сигналов Wnt / β-catenin в оральном эпителии обычно рассматривается как самый вышестоящий индуктор инициации развития зубов (Järvinen et al., 2006; Liu et al., 2008; Wang et al., 2009). Используя модели мышей in vivo, и ex vivo, , мы продемонстрировали, что стимуляция передачи сигналов Wnt / β-катенин в зубной мезенхиме имеет противоположный, то есть ингибирующий, эффект на инициацию последовательно развивающихся зубов (также известный как последовательные зубы).Генетическая активация мезенхимальной передачи сигналов Wnt / β-катенина у мышей βcat Δex3Dermo / + оказывала отрицательное влияние на последовательное формирование моляров мыши. M2 и M3 не могут развиваться во всех эксплантах зубов βcat Δex3Dermo / + , указывая тем самым, что усиление передачи сигналов мезенхимы Wnt может предотвращать зарождение зубов и что задние моляры более уязвимы для этого эффекта. С другой стороны, фармакологическая активация передачи сигналов Wnt / β-катенина ингибитором GSK BIO ингибировала последовательное развитие зубов и предотвращала образование M2 в большинстве культивируемых зачатков зубов M1 контрольных мышей.Кроме того, BIO ингибировал зарождение новых зубов в культивируемых зубных зачатках M1 эмбрионов β-cat Δex3K14 / + , которые обычно дают начало многочисленным зубам, что также указывает на то, что эти ингибирующие эффекты были вызваны стимуляцией передачи сигналов мезенхимы Wnt.
Усиленная передача сигналов Wnt / β-catenin, индуцированная BIO в эксплантатах зубов, также влияет на формирование коронки моляров, обнаруживаемой как неглубокие бугры и меньшая коронка, вероятно, в результате усиленной передачи сигналов Wnt в эпителии.Эпителиальный эффект BIO на морфогенез коронки также подтверждается наблюдением, что, как и в случае с зубными зачатками, обработанными BIO, дополнительные зубы мышей β-cat Δex3K14 / + меньше нормальных и имеют меньшее количество бугров (Järvinen et al. , 2006).
Индукция непрерывного образования зубцов принудительной эпителиальной передачей сигналов Wnt / β-катенина у мышей β-cat Δex3K14 / + (Järvinen et al., 2006) был связан с резким подавлением передачи сигналов Wnt / β-катенина в дентальной мезенхиме, о чем свидетельствует отсутствие ядерной локализации β-катенина и поразительное отсутствие репортеров Wnt / β-катенина Axin2 и Drapc1 . Механизм этого подавления, вероятно, включает индукцию растворимых эпителиальных белков, которые ингибируют передачу сигналов мезенхимы Wnt либо напрямую, либо за счет индукции ингибиторов Wnt в мезенхиме (Fig. 8). Ранее мы показали, что инициация сверхкомплектных зубов у эмбрионов β-cat Δex3K14 / + связана с индукцией многочисленных эпителиальных сигнальных центров, идентифицированных по интенсивной экспрессии β-катенина и сигнальных молекул, включая Fgf4 , Shh , Wnt10b и Bmp4 (Ярвинен и др., 2006 и не показано). Эти индуцированные сигнальные центры, вероятно, представляют IKs, ранние сигнальные центры, недавно охарактеризованные в плакодах резцов (Ahtiainen et al., 2016).
Рис. 8.
Регулирование числа зубов с помощью передачи сигналов Wnt / β-catenin. Противоположные эффекты повышенной эпителиальной и мезенхимальной передачи сигналов Wnt / β-catenin на количество зубов. Предлагаемые сигнальные сети, регулирующие передачу сигналов мезенхимального Wnt / β-catenin и предполагаемые роли Axin2 и Runx2 в этой сети, а также их мутации потери функции в генерации гиподонтии и дополнительных зубов человека, соответственно.Эпителиальные Wnts и Wnt-индуцированные эпителиальные сигнальные молекулы индуцируют ингибиторы Wnt в мезенхиме, тем самым ингибируя мезенхимальную передачу сигналов Wnt и стимулируя начало последовательного развития зубов. Ингибитор обратной связи Wnt Axin2 запускает последовательную инициацию зуба путем ингибирования передачи сигналов Wnt / β-catenin. Фактор транскрипции Runx2 в мезенхиме ингибирует ингибиторы Wnt (включая Axin2 и Drapc1) и тем самым подавляет последовательное формирование зубов. Уровень мезенхимальной передачи сигналов Wnt играет ключевую роль в инициации последовательных зубов.
Рис. 8.
Регулирование числа зубов с помощью передачи сигналов Wnt / β-catenin. Противоположные эффекты повышенной эпителиальной и мезенхимальной передачи сигналов Wnt / β-catenin на количество зубов. Предлагаемые сигнальные сети, регулирующие передачу сигналов мезенхимального Wnt / β-catenin и предполагаемые роли Axin2 и Runx2 в этой сети, а также их мутации потери функции в генерации гиподонтии и дополнительных зубов человека, соответственно. Эпителиальные Wnts и Wnt-индуцированные эпителиальные сигнальные молекулы индуцируют ингибиторы Wnt в мезенхиме, тем самым ингибируя мезенхимальную передачу сигналов Wnt и стимулируя начало последовательного развития зубов.Ингибитор обратной связи Wnt Axin2 запускает последовательную инициацию зуба путем ингибирования передачи сигналов Wnt / β-catenin. Фактор транскрипции Runx2 в мезенхиме ингибирует ингибиторы Wnt (включая Axin2 и Drapc1) и тем самым подавляет последовательное формирование зубов. Уровень мезенхимальной передачи сигналов Wnt играет ключевую роль в инициации последовательных зубов.
Теперь мы локализовали интенсивную экспрессию ингибитора Wnt Dkk4 в индуцированных IK.Dkk4 является геном-мишенью для Wnt / β-catenin (Zhang et al., 2009), и мы предполагаем, что он может диффундировать в мезенхиму, чтобы ингибировать передачу сигналов Wnt в зубах β-cat Δex3K14 / + . Расширенные домены экспрессии ингибиторов Wnt Dkk1 и Sostdc1 (также известных как Ectodin and Wise) в зубной мезенхиме, которые распространяются на мезенхиму, непосредственно лежащую в основе зубного эпителия в дополнительных зубах β-cat Δex3K14 / + , далее отражают повышенную активность эпителиального β-катенина и последующую эпителиальную передачу сигналов от IKs. Dkk1 регулируется Bmp4, Fgf4 и Shh в мезенхиме зубов и челюсти (James et al., 2006; Åberg et al., 2004; Ahn et al., 2010), а Bmp4 может индуцировать экспрессию Sostdc1 в мезенхима зачатка зуба (Laurikkala et al., 2003). Ингибиторы Wnt Dkk2 , Wif1 и Sfrp2 также были локализованы в дентальной мезенхиме и, что интересно, было показано, что они важны для последовательного формирования зубов (Jia et al., 2013, 2016).
Вероятно, что форсированная передача сигналов Wnt / β-катенина в мезенхиме зубов, как in vivo, у мышей βcat Δex3Dermo / + , так и ex vivo после введения BIO, перекрывает нормальные функции индуцированного эпителием Wnt / Ингибиторы β-катенина, таким образом нарушая важную тонкую настройку передачи сигналов мезенхимы Wnt, индуцированной эпителиальными сигналами.Обработка BIO, по-видимому, также влияет на морфогенез эпителия коронки M1, нарушая формирование и функцию вторичных EK (SEK) в эпителии. SEK экспрессируют несколько сигнальных молекул (всех консервативных путей) и другие гены, связанные с передачей сигналов, и в SEK наблюдается высокая активность Wnt (что также показано ядерным β-catenin и Lef1 в наших эксплантатах). SEK регулируют рост бугорков и, следовательно, размер коронки зуба, а небольшие SEK (и коронки зубов) в BIO-обработанных зубах указывают на негативный эффект усиления передачи сигналов Wnt / β-катенина на SEK.Это иллюстрирует сложность сигнальных сетей и контуров отрицательной обратной связи в сигнализации EK. Когда BIO был добавлен к Axin2 -null мутантным зубам в культуре, дифференцировка как мезенхимальных одонтобластов, так и эпителиальных амелобластов была ускорена (Fig. S2), дополнительно иллюстрируя множественные роли передачи сигналов Wnt во время развития зубов.
Вместе взятые, мы показали, что усиление передачи сигналов мезенхимального Wnt / β-catenin ингибирует последовательное формирование зубов, тогда как снижение передачи сигналов Wnt / β-catenin мезенхимы связано с непрерывным развитием зубов.Эти результаты показывают, что эпителиальная передача сигналов Wnt / β-катенина, которая инициирует формирование зубов и индуцирует развитие дополнительных зубов, подавляет активность передачи сигналов Wnt / β-катенина в мезенхиме посредством прямой, а также косвенной стимуляции ингибиторов Wnt. Мы заключаем, что уровень передачи сигналов мезенхимы Wnt является критическим для последовательной инициации зубов (Рис. 8).
«Модель ингибирующего каскада» была построена на основе ex vivo анализа эффектов сигнальных молекул на последовательное образование моляров мыши (Kavanagh et al., 2007). Согласно этой модели, модуляция баланса активаторов и ингибиторов определяет размеры M2 и M3. В частности, усиление ингибирования имеет кумулятивный эффект на более задние зубы, а размер более раннего моляра определяет, будет ли формироваться следующий моляр или нет. Наши результаты показывают, что передача сигналов мезенхимального Wnt / β-катенина является ингибитором в этой модели, поскольку усиление передачи сигналов мезенхимального Wnt / β-катенина в мутантах βcat Δex3Dermo / + не предотвращало развитие M1, но подавляло образование M2. и М3.Хотя усиление передачи сигналов β-catenin, индуцированное BIO, не нарушает драматически продолжающееся развитие M1, вполне вероятно, что это влияет на передачу сигналов от M1 к M2. Взятые вместе, мы интерпретируем наши результаты как указание на ингибирующую роль передачи сигналов мезенхимы Wnt в последовательном формировании зубов.
Существует несколько моделей мышей, в которых модуляция ключевых сигнальных путей ставит под угрозу последовательное формирование моляров.Классическим примером являются мыши Eda-null Tabby , у которых примерно в половине случаев отсутствует один или несколько M3 (Grüneberg, 1966). Eda является мишенью для передачи сигналов Wnt / β-catenin в эпителии и регулирует др. Пути передачи сигналов, включая Shh, Fgf и Bmp в зубах (Lefebvre and Mikkola, 2014). Мишень Eda в зубном эпителии, Fgf20 , была указана как важный активатор образования зубов в модели ингибирующего каскада (Häärä et al., 2012). Кроме того, снижение дозы Pax9 , мезенхимального фактора транскрипции, регулируемого передачей сигналов Fgf (Neubüser et al., 1997), прогрессивно снижает количество моляров: небольшое снижение экспрессии Pax9 ингибирует только развитие M3, более высокое снижение ингибирует как M3, так и M2, а полная делеция функции Pax9 предотвращает образование всех трех моляров (Kist et al. др., 2005). Во всех этих примерах подавление образования заднего моляра является результатом ингибирования консервативных сигнальных путей. Однако наши результаты показывают, что стимуляция сигнальных путей, в частности Wnt и Bmp, также может ингибировать последовательное формирование зубов.
Передача сигналов Wnt / β-catenin и Bmp тесно связана на всех стадиях формирования зубов и они влияют как на эпителиальные, так и мезенхимальные ткани (O’Connell et al., 2012). Перекрестное взаимодействие путей Wnt и Bmp в дентальной мезенхиме является критическим для раннего морфогенеза моляров мышей и их последовательного образования (Jia et al., 2013). В резце мезенхимальная делеция β-катенина привела к подавлению экспрессии Bmp4 , что нарушило целостность эпителиальной зубной плакоды и привело к развитию двух тонких резцов (Fujimori et al., 2010). Мы наблюдали, что ингибирование образования M2 ex vivo с помощью BIO приводит к усиленной экспрессии Bmp4 в мезенхиме зубов и его гена-мишени Id1 в эпителии зубов. Кроме того, Bmp4 предотвращал образование M2 аналогично BIO. Однако ингибитор Bmp Noggin не устранял BIO-индуцированное ингибирование развития M2, указывая на то, что эффект усиленной передачи сигналов Wnt лишь частично опосредован Bmp.
Wnt / β-catenin, по-видимому, является негативным регулятором передачи сигналов Fgf в дентальной мезенхиме. Экспрессия Fgf10 умеренно подавлялась в BIO-обработанных зубах, но неожиданно была повышена регуляция репортера Fgf Dusp4 . Возможное объяснение этого — компенсация Fgf2 и / или Fgf3, которые также были активированы BIO. Ранее мы продемонстрировали, что BIO ингибирует экспрессию Fgf10 в зубной мезенхиме, окружающей шейную петлю непрерывно растущего резца (Yang et al., 2015). Это привело к апоптозу эпителиальных стволовых клеток в петле шейки матки, и в этом случае действие BIO могло быть устранено с помощью Fgf10.Таким образом, передача сигналов мезенхимального Wnt / β-катенина стимулирует экспрессию Bmp4 и ингибирует экспрессию Fgf10 как в резцах, так и в молярах.
У людей гетерозиготные мутации потери функции в AXIN2 , гене-мишени прямого Wnt и ингибиторе обратной связи, вызывают различную степень гиподонтии (Lammi et al., 2004; Marvin et al., 2011; Wong et al., 2014; Юэ и др., 2016). Наши результаты предполагают, что этот фенотип является результатом усиленной передачи сигналов Wnt / β-catenin мезенхимы. Подобные фенотипы также вызваны мутациями в гене WNT10A , которые составляют более 30% случаев агенеза зубов с известными мутациями (van den Boogaard et al., 2012; Arte et al., 2013; Arzoo et al., 2014; Mues et al., 2014; Yin and Bian, 2015), что указывает на ключевую роль Wnt10a в развитии зубов. Во время инициации развития зубов у мышей экспрессия Wnt10a, и других лигандов Wnt, а также сигнальная активность Wnt / β-катенина в основном ограничиваются эпителием (Dassule and McMahon, 1998; Liu et al., 2008), а стабилизация передачи сигналов β-catenin в оральном эпителии эмбрионов мышей ведет к непрерывному зарождению новых зубов (Järvinen et al., 2006). Мы демонстрируем, что это ведет к драматическому подавлению активности Axin2 в мезенхиме и усилению инициации зубов, подтверждая, что эпителиальные лиганды Wnt функционируют выше Axin2 (Figs 1F и 8).
Паттерн экспрессии Axin2 во время последовательного образования моляров мыши указывает на сигнальную активность Wnt в мезенхиме, лежащей в основе заднего хвоста M1, или континуальной ламине, где инициируется M2.Мы предполагаем, что усиление передачи сигналов β-catenin в мезенхиме в этом сайте ингибирует образование M2. У человека молочные зубы отсутствуют редко, и чаще всего отсутствуют постоянные зубы, которые развиваются последними в пределах класса зубов, включая третий моляр (M3), второй верхний резец (I2) и второй премоляр (P2). Эти зубы также чаще всего поражаются агенезом зубов, вызванным мутациями AXIN2 и WNT10A (Lammi et al., 2004; Arte et al., 2013; Arzoo et al., 2014; Ян и др., 2015). С другой стороны, гетерозиготные мутации с потерей функции в RUNX2 при синдроме кледокраниальной дисплазии человека стимулируют последовательное развитие зубов, что приводит к чрезмерному образованию замещающих зубов и дополнительных задних моляров (Jensen and Kreiborg, 1990; Lee et al., 1997 ). Поскольку замена зуба и добавление моляра представляют собой сопоставимые процессы развития, характеризующиеся последовательным инициированием зубной пластинки, связанной с ранее сформированным зубом (Juuri et al., 2013), мы предполагаем, что мезенхимальный β-катенин играет сходную ингибирующую роль в обоих процессах.
Насколько нам известно, образование лишних зубов, вызванное мутациями RUNX2 , ранее не было связано с передачей сигналов Wnt. Мы обнаружили коэкспрессию Runx2 и Axin2 в зубной мезенхиме мышей во время стадии зачатка, а также в ассоциации с инициацией M2, предвосхищая механизмы реципрокной регуляции между этими генами.Runx2 важен для передачи сигналов Fgf от зубного эпителия к мезенхиме на ранних стадиях развития зубов (Åberg et al., 2004). Однако, поскольку задержка развития на стадии зачатка Runx2- нулевых мутантных зубов не восстанавливается ни с помощью Fgf, ни с помощью Shh, мы предположили, что задействованы др. Сигнальные пути (Åberg et al., 2004). Мы предполагаем, что Runx2 обеспечивает передачу сигналов Wnt во время почкования первых зубов, а также во время последовательного формирования зубов. Предыдущие исследования показали связь между передачей сигналов Runx2 и Wnt во время развития костей черепа и продемонстрировали, что Runx2 регулирует репрессию Axin2 . Axin2 активируется в Runx2-дефицитных мезенхимальных клетках, а Runx2 непосредственно репрессирует Axin2 (Li et al., 2009; McGee-Lawrence et al., 2013). В соответствии с этими исследованиями, наш анализ продемонстрировал, что экспрессия Axin2 , а также др. Ингибитора обратной связи Wnt Drapc1 активируется в зачатках нулевых мутантных зубов Runx2- в мезенхиме, окружающей развивающийся моляр мыши.
Интересно, что стоматологические фенотипы Axin2 и Runx2 различаются у человека и мыши.Отсутствие каких-либо дефектов зубов у мышей Axin2 -null, которые, как сообщается, демонстрируют нормальный общий фенотип только с легким дефектом костей черепа (Yu et al., 2005), демонстрирует существенное различие между человеком и мышью в чувствительности к снижению функции Axin2. . Вероятно, что у мышей функция Axin2 компенсируется еще неизвестными регуляторными механизмами передачи сигналов Wnt / β-катенина, например Drapc1 , другой ингибитор обратной связи передачи сигналов Wnt / β-catenin, который обнаруживает коэкспрессию с Axin2 в зубных зачатках мыши.Одновременное увеличение экспрессии Axin2 и Drapc1 у нулевых мутантов Runx2- подтверждает наличие компенсаторной сигнальной петли между этими двумя генами, которая потенциально может объяснять отсутствие фенотипа зубов у нулевых мышей Axin2-. Можно предположить, что у людей может быть меньше компенсации другим ингибитором (ами) Wnt, что объясняет уменьшение числа зубов у гетерозигот человека AXIN2 . Кроме того, в отличие от человека с фенотипом гипердонтии RUNX2 +/- , гетерозиготные мыши Runx2 не имеют фенотипа зубов, а у нулевых мутантных мышей Runx2- отсутствуют все зубы из-за остановки их развития на стадии зачатка. (Д’Суза и др., 1999). Стимуляция последовательного развития зубов у гетерозигот RUNX2 человека не может быть связана с задержкой развития M1 у нулевых мышей Runx2-. Отмеченные различия могут быть результатом различий в составе и функциях зубных рядов человека и мыши.
Взятые вместе, наши результаты показывают, что мезенхимальная передача сигналов Wnt / β-catenin является основным негативным регулятором последовательного формирования зубов и что она ограничивает количество зубов.Мы предполагаем, что непрерывное последовательное развитие лишних зубов, вызванное гетерозиготными мутациями потери функции в гене RUNX2 человека, происходит из-за отсутствия репрессии AXIN2 и других ингибиторов Wnt, таких как DRAPC1 с помощью RUNX2, что приводит к снижению передачи сигналов мезенхимы Wnt. Также есть соблазн предположить, что модуляция передачи сигналов мезенхимального Wnt / β-catenin с помощью Runx2 и Wnt ингибиторов, включая Axin2 и Drapc1, возможно, сыграла важную роль в эволюции, когда потенциал для непрерывной инициации зубов был утрачен у большинства млекопитающих.
Генерация мышей β-cat Δex3K14 / + была описана ранее (Järvinen et al., 2006). Мыши K14-cre были подарком Макото М. Такето (Университет Киото, Япония). Мыши с мутантами β-catenin- flox-ex3 и Axin2 -null (Yu et al., 2005) были подарками Walter Birchmeier (Центр Макса Дельбрука, Берлин, Германия). βcat Δex3Dermo / + эмбрионов были получены путем скрещивания мышей Dermo-cre ( Twist2-cre ) с мышами βcat -flox-ex3 (Harada et al., 1999). Мыши Dermo-cre были подарком Кристины Хартманн (Исследовательский институт молекулярной патологии, Вена, Австрия). Нуль-мутантные мыши Runx2- были подарком Майкла Оуэна (Imperial Cancer Research Fund, Лондон, Великобритания) (Mundlos et al., 1997; Åberg et al., 2004). Генерация Lef1 -репортерных мышей (TCF / Lef: h3B-GFP) была описана в другом месте (Ferrer-Vaquer et al., 2010). Голых мышей с ослабленным иммунитетом (лаборатория Джексона) использовали в экспериментах по трансплантации капсулы почки.
Собранные ткани фиксировали в течение ночи в 4% параформальдегиде, обезвоживали и заливали парафином. Срезы в фронтальной и сагиттальной плоскости разрезали на 5-7 мкм и обрабатывали для гистологического анализа, иммунофлуоресценции и радиоактивной гибридизации in situ и . Передача сигналов активированного Wnt анализировалась с помощью иммунофлуоресценции на срезах, обработанных 10 мМ натрийцитратным буфером (pH 6.0) для поиска антигена с последующей инкубацией с антителом против β-катенина мыши (BD Pharmingen) в разведении 1: 1000 и вторичным антителом против AF568 (Invitrogen) в разведении 1: 500.
Radioactive in situ гибридизацию проводили с использованием стандартного протокола (Wilkinson and Green, 1990). Меченые [ 35 S] -UTP (PerkinElmer) РНК-зонды для следующих генов использовали для обнаружения экспрессии генов: Axin2 (Lammi et al., 2004), Drapc1 (Jukkola et al., 2004), Dkk1 (James et al., 2006), Sostdc1 (Laurikkala et al., 2003), Shh (Vaahtokari et al., 1996 ), Fgf4 (Jernvall et al., 1994), Lef1 (Travis et al., 1991), Fgf3 (Järvinen et al., 2006), Runx2 (James et al., 2006), Fgf 10 (Yang et al., 2015), Dusp6 (James et al., 2006), Id1 (Rice et al., 2000), Bmp4 (Vaahtokari et al., 1996) и Dkk4 (Fliniaux et al., 2008).
Обозначение стоматологической анатомии | Bücco
Эта статья представляет собой публикацию о здоровье полости рта, выпущенную Орденом стоматологов Квебека.
Арт. и © Орден дантистов Квебека
Экстремальное увеличение зубов до атомарного уровня!
Хотя эмаль известна как самая твердая ткань человеческого тела и, несмотря на то, что она кажется гладкой, а не пористой, оценка с помощью электронного микроскопа дает нам совершенно иное восприятие этой ткани.Это исключительное видео показывает, как эмаль, состоящая из крохотных элементов, называемых призмами, на самом деле имеет очень неровную поверхность при увеличении в тысячи и миллионы раз. Специальные клетки производят эмаль только один раз при формировании зубов; эмаль не подлежит замене, если она изношена или повреждена.
Посмотрите это видео, чтобы узнать больше… изображения говорят сами за себя!
➡
Чтобы узнать больше о нормальном прорезывании зубов и связанных с ними аномалиях.
Стоматологическая запись — нумерация зубов
Чтобы говорить на одном языке между стоматологами и стоматологическим персоналом, мы используем систему, которая позволяет нам точно идентифицировать каждый зуб по номеру. Это предотвращает путаницу и сводит к минимуму вероятность ошибки в письменном или устном общении.
Международная федерация стоматологии
- Чаще всего используется система Fédération Dentaire Internationale (FDI), которая используется в Квебеке, Канаде и большинстве европейских стран.
- Каждый зуб представлен двузначным числом. Каждая верхняя и нижняя дуги содержат по 8 постоянных зубов, пронумерованных от средней линии к спине от 1 до 8 для зубного ряда взрослого.
- Каждая арка разделена на 2 квадрантов или полуарок, правую и левую, обозначенных первой цифрой. Правый верхний квадрант — №1, а левый — №2. Нижняя арка включает квадранты № 3 (внизу слева) и № 4 (внизу справа).
- Зубы в квадранте № 1 пронумерованы, начиная с «1», за которым следует номер, идентифицирующий зуб (от 1 до 8).Зубы в квадранте № 2 начинаются с цифры «2» и так далее для каждого квадранта.
- В каждом квадранте 8 постоянных зубов, пронумерованных следующим образом:
- Резцы : верхний правый (11, 12), верхний левый (21, 22), нижний левый (31, 32) и нижний правый (41, 42) (всего 8 резцов).
- Клыки : верхний правый (13), верхний левый (23), нижний левый (33) и нижний правый (43) (4 клыка).
- Премоляры : верхний правый (14, 15), верхний левый (24, 25), нижний левый (34, 35) и нижний правый (44, 45) (всего 8 премоляров).
- Моляры : верхний правый (16, 17), верхний левый (26, 27), нижний левый (36, 37) и нижний правый (46, 47).
- Зубы мудрости (третьи моляры): верхний правый (18), верхний левый (28), нижний левый (38) и нижний правый (48) (всего 12 коренных зубов).
- (Приведенная выше иллюстрация воспроизведена и изменена с разрешения автора.)
- Следует отметить, что «Правый» и «Левый» относятся к правой и левой стороне пациента, когда мы смотрим на него перед собой.
Зубные поверхности
Причем у каждого зуба несколько поверхностей:
- Labial : поверхность со стороны губ передних зубов (резцов и клыков).
- Буккальный : поверхность со стороны щек задних зубов (премоляры и коренные зубы).
- Lingual : поверхность со стороны языка каждого зуба (переднего и заднего).
- Окклюзионный : жевательная поверхность боковых зубов.
- Incisive : острый край резцов.
- Mésial : межзубная поверхность (между соседними зубами), ближайшая к средней линии.
- Дистальный : межзубная поверхность (между соседними зубами), наиболее удаленная от средней линии.
Универсальная стоматологическая система нумерации (США)
- Эта система нумерации, в основном используемая в США, начинается с нумерации зубов цифрой 1 от верхней правой части (третий моляр) и заканчивается на нижней правой челюсти зубом № 32 (третий моляр).
- Таким образом, наш зуб № 18 — это зуб № 1 для американцев, зуб № 11 — их зуб № 9, наш зуб № 26 — их № 15, наш № 38 — их № 19, наш № 31 — их № 26, наш №. 48 — это их №36 и т. Д.
- Вы можете представить себе потенциальную путаницу, если бы для связи между стоматологами, использующими разные системы, использовались только номера зубов. Вот почему стоматологическая диаграмма (одонтограмма), показывающая, какие зубы мы имеем в виду, часто включается в описание зубных рядов в рецепты стоматологических работ (удаление, реставрация, операции и т. Д.)).
Изображение различных частей и компонентов зуба (нижний резец) и пародонта (альвеолярная кость и ткань десны).
Морфология небных складок связана с вариациями номера зуба
Standring, S. Анатомия Грея: академические основы клинической практики 41-е изд. (Эльзевир, Амстердам, 2016).
Google ученый
Hauser, G., Daponte, A. & Roberts, M. J. Небные складки. J. Anat. 165 , 237–249 (1989).
CAS Google ученый
Петеркова Р., Клепакек И. и Петерка М. Пренатальное развитие небных складок у мышей: сканирующая электронная микроскопия и гистологические исследования. J. Craniofac. Genet. Dev. Биол. 7 , 169–189 (1987).
CAS Google ученый
Итикава, Х., Мацуо, С., Силос-Сантьяго, И., Жакин, М. Ф. и Сугимото, Т. Зависимость развития окончаний Меркель от мышц неба. Brain Res. Мол. Brain Res. 88 , 171–175 (2001).
CAS Статья Google ученый
Nunzi, M. G., Pisarek, A. & Mugnaini, E. Клетки Меркеля, корпускулярные нервные окончания и свободные нервные окончания в слизистой оболочке неба мыши экспрессируют три подтипа везикулярных переносчиков глутамата. J. Neurocytol. 33 , 359–376. https://doi.org/10.1023/B:NEUR.0000044196.45602.92 (2004).
CAS Статья Google ученый
Капали С., Таунсенд Г., Ричардс Л. и Пэриш Т. Образцы небных складок у австралийских аборигенов и кавказцев. Aust. Вмятина. J. 42 , 129–133 (1997).
CAS Статья Google ученый
Lysell, L. Plicae palatinae transversae and papilla incisiva у человека; морфологическое и генетическое исследование. Acta Odontol. Сканд. 13 , 5–137 (1955).
CAS Google ученый
Мердок, А. М., Патир, А., Сеймен, Ф. и Виейра, А. Р. Исследования небных складок и вариаций фактора регуляции интерферона 6 в группе семей со спорадической гиподонтией. J. Oral. Sci. 51 , 521–526 (2009).
CAS Статья Google ученый
Пиви Д. К. мл. И Кендрик Г. С. Влияние движения зубов на небные складки. J. Prosthet. Вмятина. 18 , 536–542 (1967).
Артикул Google ученый
Thomas, C.J. & Kotze, T.J. Рисунок небной складки: новая классификация. J. Dent. Доц. S. Afr. 38 , 153–157 (1983).
CAS Google ученый
Christou, P. & Kiliaridis, S. Связанные с вертикальным ростом изменения положения небных складок и резцов верхней челюсти. Am. J. Orthod. Дентофак. Ортоп. 133 , 81–86. https://doi.org/10.1016/j.ajodo.2007.07.009 (2008).
Артикул Google ученый
Ким, Х. К., Мун, С. К., Ли, С. Дж.И Парк Ю.С. Трехмерное биометрическое исследование небных складок у детей с анализом смешанной модели: 9-летнее продольное исследование. Am. J. Orthod. Дентофак. Ортоп. 141 , 590–597. https://doi.org/10.1016/j.ajodo.2011.11.018 (2012).
Артикул Google ученый
Byatnal, A. et al. Палатоскопия: дополнение к судебной одонтологии — сравнительное исследование пяти различных популяций Индии. J. Nat. Sci. Биол. Med. 5 , 52–55. https://doi.org/10.4103/0976-9668.127287 (2014).
Артикул Google ученый
Де Анжелис, Д., Риболи, Ф., Гибелли, Д., Каппелла, А. и Каттанео, С. Небные складки как индивидуализирующий маркер: надежность для судебной стоматологии и идентификации личности. Sci. Правосудие 52 , 181–184. https://doi.org/10.1016/j.scijus.2011.09.002 (2012).
Артикул Google ученый
Джайн А. и Чоудхари Р. Небные складки и их роль в судебной стоматологии. J. Investig. Clin. Вмятина. 5 , 171–178. https://doi.org/10.1111/j.2041-1626.2013.00150.x (2014).
Артикул Google ученый
Томас, К. Дж. И Коце, Т. Дж. Небные складки в судебной стоматологии. J. Forensic Odontostomatol. 1 , 11–18 (1983).
CAS Google ученый
Jiang, T. X., Jung, H. S., Widelitz, R. B. & Chuong, C. M. Самоорганизация периодических паттернов диссоциированными мезенхимальными клетками пера и регуляция размера, количества и расстояния между зачатками. Разработка 126 , 4997–5009 (1999).
CAS Google ученый
Кондо С. и Асал Р. Реакционно-диффузионная волна на коже морского ангела Pomacanthus. Nature 376 , 765–768 (1995).
ADS CAS Статья Google ученый
Моу, К., Джексон, Б., Шнайдер, П., Овербек, П. А. и Хедон, Д. Дж. Генерация первичного рисунка волосяных фолликулов. Proc. Natl. Акад. Sci. США 103 , 9075–9080. https://doi.org/10.1073 / pnas.0600825103 (2006).
ADS CAS Статья Google ученый
Sick, S., Reinker, S., Timmer, J. & Schlake, T. WNT и DKK определяют расстояние между волосяными фолликулами посредством механизма реакции-диффузии. Наука 314 , 1447–1450. https://doi.org/10.1126/science.1130088 (2006).
ADS CAS Статья Google ученый
Тьюринг, А. М. Химические основы морфогенеза: модель реакции-диффузии для развития. Philos. Пер. R. Soc. Лондон. 237 , 37–72 (1952).
ADS МАТЕМАТИКА Google ученый
Грин Дж. Б. и Шарп Дж. Позиционная информация и реакция – диффузия: объединяются две большие идеи в биологии развития. Развитие 142 , 1203–1211. https://doi.org/10.1242/dev.114991 (2015).
CAS Статья Google ученый
Economou, A. D. et al. Периодическое образование полосок по механизму Тьюринга, действующему в зонах роста на нёбе млекопитающих. Nat. Genet. 44 , 348–351. https://doi.org/10.1038/ng.1090 (2012).
CAS Статья Google ученый
Pantalacci, S. et al. Построение рисунка небных складок путем последовательного добавления позволяет выявить переднюю / заднюю границу небного развития. BMC Dev. Биол. 8 , 116. https://doi.org/10.1186/1471-213X-8-116 (2008).
CAS Статья Google ученый
Welsh, I.C. & O’Brien, T.P. Интеграция сигналов в зоне роста морщин направляет формирование последовательного сигнального центра SHH во время рострального отростка неба. Dev. Биол. 336 , 53–67. https://doi.org/10.1016/j.ydbio.2009.09.028 (2009 г.).
CAS Статья Google ученый
Lin, C. et al. Индуктивная роль передачи сигналов Wnt-beta-Catenin в формировании орального аппарата. Dev. Биол. 356 , 40–50. https://doi.org/10.1016/j.ydbio.2011.05.002 (2011).
CAS Статья Google ученый
Zhang, Z. et al. Спасение волчьей пасти у Msx1-дефицитных мышей с помощью трансгенного Bmp4 выявляет сеть передачи сигналов BMP и Shh в регуляции палатогенеза у млекопитающих. Разработка 129 , 4135–4146 (2002).
CAS Google ученый
Trakanant, S. et al. Молекулярные механизмы в развитии небных складок. J. Oral Biosci. https://doi.org/10.1016/j.job.2019.12.002 (2019).
Артикул Google ученый
Кобурн, М. Т. и Грин, Дж. Б. Передача сигналов ежа в развитии вторичного неба. Фронт. Oral Biol. 16 , 52–59. https://doi.org/10.1159/000337543 (2012).
Артикул Google ученый
Кобурн, М. Т. и Шарп, П. Т. Составление чисел: молекулярный контроль зубной формулы млекопитающих. Семин. Cell Dev. Биол. 21 , 314–324. https://doi.org/10.1016/j.semcdb.2010.01.007 (2010).
CAS Статья Google ученый
He, F. & Chen, Y. Передача сигналов Wnt в развитии губ и неба. Фронт. Oral Biol. 16 , 81–90. https://doi.org/10.1159/000337619 (2012).
Артикул Google ученый
Parada, C. & Chai, Y. Роль сигнального пути BMP в развитии губ и неба. Фронт. Oral Biol. 16 , 60–70. https://doi.org/10.1159/000337617 (2012).
Артикул Google ученый
Stanier, P. & Pauws, E. Развитие губы и неба: передача сигналов FGF. Фронт. Oral Biol. 16 , 71–80. https://doi.org/10.1159/000337618 (2012).
Артикул Google ученый
Vastardis, H., Karimbux, N., Guthua, S. W., Seidman, J. G. & Seidman, C. E. Миссенс-мутация гомеодомена MSX1 человека вызывает селективный агенез зубов. Nat. Genet. 13 , 417–421. https://doi.org/10.1038/ng0896-417 (1996).
CAS Статья Google ученый
Stockton, D. W., Das, P., Goldenberg, M., D’Souza, R. N. & Patel, P. I. Мутация PAX9 связана с олигодонтией. Nat. Genet. 24 , 18–19. https://doi.org/10.1038/71634 (2000).
CAS Статья Google ученый
Kantaputra, P. & Sripathomsawat, W. WNT10A и изолированная гиподонтия. Am. J. Med. Genet. А 155А , 1119–1122. https://doi.org/10.1002/ajmg.a.33840 (2011 г.).
CAS Статья Google ученый
Lammi, L. et al. Мутации в AXIN2 вызывают семейную агенезию зубов и предрасполагают к колоректальному раку. Am. J. Hum. Genet. 74 , 1043–1050. https://doi.org/10.1086/386293 (2004).
CAS Статья Google ученый
Kawasaki, M. et al. Lrp4 / Wise регулирует развитие небных складок посредством механизмов реакции-диффузии типа Тьюринга. PLoS ONE 13 , e0204126.https://doi.org/10.1371/journal.pone.0204126 (2018).
CAS Статья Google ученый
Silva-Sousa, A.C. et al. Лево-правая асимметрия небных складок ассоциирована с генетическими вариантами пути передачи сигналов WNT. Arch. Oral Biol. 110 , 104604. https://doi.org/10.1016/j.archoralbio.2019.104604 (2019).
CAS Статья Google ученый
Моран А., Типпет Х., Манохаран А. и Кобурн М. Т. Изменение рисунка небных складок у субъектов с олигодонтией: пилотное исследование. Am. J. Orthod. Дентофак. Ортоп. 150 , 295–302. https://doi.org/10.1016/j.ajodo.2015.12.030 (2016).
Артикул Google ученый
Fournier, B.P. et al. Паттерны агенеза зубов подчеркивают природу причинных мутировавших генов. J. Dent. Res. 97 , 1306–1316. https://doi.org/10.1177/0022034518777460 (2018).
CAS Статья Google ученый
Кук, Р. Дж. И Фэруэлл, В. Т. Соображения множественности при разработке и анализе клинических испытаний. J. R. Stat. Soc. 159 , 93–110 (1996).
Артикул Google ученый
Ибечу, П. К., Дидия, Б. К. и Аригбеде, А. О. Сравнительное исследование структуры небных складок у этнических групп игбо и икверре в Нигерии: исследование университета Порт-Харкорта. Анат. Res. Int. 2014 , 123925. https://doi.org/10.1155/2014/123925 (2014).
CAS Статья Google ученый
Surekha, R. et al. Оценка структуры небных складок у населения Манипури и Кералы. J. Forensic Dent. Sci. 4 , 93–96. https://doi.org/10.4103/0975-1475.109896 (2012).
CAS Статья Google ученый
Hauser, G. & Roberts, M. J. Небные складки в Свази, южная Африка. Антрополь. Anz. 44 , 257–261 (1986).
CAS Google ученый
Шетти, С. К., Калиа, С., Патил, К.И Махима, В. Г. Рисунок небных складок у жителей Майсора и Тибета. Индиан Дж. Дент. Res. 16 , 51–55 (2005).
Google ученый
Бабу, Г. С., Бхарат, Т. С. и Кумар, Н. Г. Характеристики рисунка небных складок у населения западного годавари в Индии. J. Clin. Диаг. Res. 7 , 2356–2359. https://doi.org/10.7860/JCDR/2013/6058.3523 (2013).
Артикул Google ученый
Gondivkar, S. M. et al. Морфологическое исследование небных складок у населения Западной Индии. J. Forensic Leg. Med. 18 , 310–312. https://doi.org/10.1016/j.jflm.2011.06.007 (2011).
Артикул Google ученый
Раджан, В. П., Джон, Дж. Б., Сталин, А., Прия, Г. и Абутагир, А. К. Морфология рисунков небных складок у детей 5–15 лет. J. Pharm. Bioallied Sci. 5 , С43-47. https://doi.org/10.4103/0975-7406.113295 (2013).
Артикул Google ученый
Saxena, E. et al. Исследование структуры небных складок у мужчин, женщин и трансгендеров в городе Бхопал. J. Forensic Dent. Sci. 7 , 142–147. https://doi.org/10.4103/0975-1475.146370 (2015).
Артикул Google ученый
Бхагват, С. и Чандра, Л. Образец Руга в выборке населения Мирута: институциональное исследование. J. Forensic Dent. Sci. 6 , 122–125. https://doi.org/10.4103/0975-1475.132542 (2014).
CAS Статья Google ученый
Бхарат, С. Т., Кумар, Г. Р., Дханапал, Р. и Сарасвати, Т. Определение пола с помощью анализа дискриминантной функции небных складок из популяции прибрежной Андхры. J. Forensic Dent. Sci. 3 , 58–62. https://doi.org/10.4103/0975-1475.
Артикул Google ученый
Сараф, А., Бедиа, С., Индуркар, А., Дегвекар, С. и Бховате, Р. Паттерны Руга как дополнение к дифференциации пола при судебно-медицинской идентификации. J. Forensic Odontostomatol. 29 , 14–19 (2011).
CAS Google ученый
van der Linden, F. P. Изменения положения боковых зубов по отношению к остриям морщин. Am. J. Orthod. 74 , 142–161 (1978).
Артикул Google ученый
Hosokawa, R. et al. Эпителиально-специфическая потребность в передаче сигналов FGFR2 во время развития зубов и неба. J. Exp. Zool. B Mol. Dev. Evol. 312B , 343–350. https://doi.org/10.1002/jez.b.21274 (2009 г.).
CAS Статья Google ученый
Welsh, I.C., Hagge-Greenberg, A. & O’Brien, T.P. Зависимая от дозировки роль Spry2 в росте и формировании паттерна во время развития неба. мех. Dev. 124 , 746–761. https://doi.org/10.1016/j.mod.2007.06.007 (2007).
CAS Статья Google ученый
Lee, J. M. et al. Передача сигналов Shh важна для морфогенеза морфогенеза у мышей. Histochem. Cell Biol. 136 , 663–675. https://doi.org/10.1007/s00418-011-0870-7 (2011).
CAS Статья Google ученый
Khalaf, K., Miskelly, J., Voge, E. & Macfarlane, T. V. Распространенность гиподонтии и связанных факторов: систематический обзор и метаанализ. J. Orthod. 41 , 299–316. https: // doi.org / 10.1179 / 1465313314Y.0000000116 (2014).
Артикул Google ученый
Кобурн, М. Т. и Шарп, П. Т. Заболевания зубов: генетические и молекулярные основы наследственных аномалий, влияющих на зубной ряд. Wiley междисциплинарный. Rev. Dev. Биол. 2 , 183–212. https://doi.org/10.1002/wdev.66 (2013).
Артикул Google ученый
Гликозаминогликаны, катализируемые FAM20B, контролируют количество зубов мышей путем ограничения передачи сигналов FGFR2b | BMC Biology
Вайнио С., Линь Ю. Органогенез: координация раннего развития почек: уроки нацеливания на гены. Nat Rev Genet. 2002; 3 (7): 533.
CAS PubMed Google ученый
Balic A, Thesleff I. Взаимодействия тканей, регулирующие развитие и обновление зубов. Curr Top Dev Biol. 2015; 115: 157–86.
PubMed Google ученый
Thesleff I, Barrach HJ, Foidart JM, Vaheri A, Pratt RM, Martin GR.Изменения в распределении коллагена IV типа, ламинина, протеогликана и фибронектина во время развития зубов у мышей. Dev Biol. 1981; 81 (1): 182–92.
CAS PubMed Google ученый
Теслефф И., Хурмеринта К. Взаимодействие тканей в развитии зубов. Дифференциация. 1981. 18 (1–3): 75–88.
CAS PubMed Google ученый
Thesleff I, Vaahtokari A, Partanen AM.Регуляция органогенеза. Общие молекулярные механизмы, регулирующие развитие зубов и других органов. Int J Dev Biol. 1995. 39 (1): 35–50.
CAS PubMed Google ученый
Bai XM, Van der Schueren B., Cassiman JJ, Van den Berghe H, David G. Дифференциальная экспрессия протеогликанов гепарансульфата на множественной клеточной поверхности во время эмбрионального развития зубов. J Histochem Cytochem. 1994. 42 (8): 1043–54.
CAS PubMed Google ученый
Wu J, Li H, Han L, Sun T, Tian Y, Wang X. Пространственно-временная картина экспрессии Syndecans в эмбриональных зубах мыши. Паттерны экспрессии генов 2020; 36: 119109. https://doi.org/10.1016/j.gep.2020.119109.
Mythreye K, Blobe GC. Корецепторы передачи сигналов протеогликана: роль в клеточной адгезии, миграции и инвазии. Сотовый сигнал. 2009. 21 (11): 1548–58.
Коике Т., Изумикава Т., Тамура Дж. И., Китагава Х. FAM20B представляет собой киназу, которая фосфорилирует ксилозу в области гликозаминогликана-белковой связи.Биохим Дж. 2009; 421 (2): 157–62.
CAS PubMed Google ученый
Вэнь Дж., Сяо Дж., Рахдар М., Чоудхури Б.П., Цуй Дж., Тейлор Г.С., Эско Дж. Д., Диксон Дж. Фосфорилирование ксилозы функционирует как молекулярный переключатель, регулирующий биосинтез протеогликанов. P Natl Acad Sci USA. 2014. 111 (44): 15723–8.
CAS Google ученый
Фогель П., Хансен Г.М., Рид РВ, Вэнс Р.Б., Тиль М., Лю Дж., Вронски Т.Дж., Смит Д.Д., Джетер-Джонс С., Броммейдж Р.Несовершенный амелогенез и другие дефекты биоминерализации у мышей без Fam20a и Fam20c. Vet Pathol. 2012. 49 (6): 998–1017.
CAS PubMed Google ученый
Tian Y, Ma P, Liu C, Yang X, Crawford DM, Yan W, Bai D, Qin C, Wang X. Инактивация Fam20B в зубном эпителии мышей приводит к появлению лишних резцов. Eur J Oral Sci. 2015; 123 (6): 396–402.
CAS PubMed PubMed Central Google ученый
Thesleff I, Jalkanen M, Vainio S, Bernfield M. Экспрессия протеогликанов на клеточной поверхности коррелирует с эпителиально-мезенхимальным взаимодействием во время морфогенеза зубов. Dev Biol. 1988. 129 (2): 565–72.
CAS PubMed Google ученый
Вайнио С., Ялканен М., Теслефф И. Экспрессия синдекана и тенасцина индуцируется эпителиально-мезенхимальными взаимодействиями в мезенхиме эмбриональных зубов. J Cell Biol. 1989. 108 (5): 1945–53.
CAS PubMed Google ученый
Salmivirta M, Elenius K, Vainio S, Hofer U, Chiquet-Ehrismann R, Thesleff I, Jalkanen M. Синдекан из мезенхимы эмбрионального зуба связывает тенасцин. J Biol Chem. 1991. 266 (12): 7733–9.
CAS PubMed Google ученый
Vainio S, Jalkanen M, Vaahtokari A, Sahlberg C, Mali M, Bernfield M, Thesleff I. Экспрессия гена синдекана индуцируется на ранней стадии, носит временный характер и коррелирует с изменениями в пролиферации мезенхимальных клеток во время органогенеза зубов.Dev Biol. 1991. 147 (2): 322–33.
CAS PubMed Google ученый
Мизумото С., Ямада С., Сугахара К. Генетические нарушения человека и нокаутные мыши с дефицитом гликозаминогликана. Biomed Res Int. 2014; 2014: 495764.
PubMed PubMed Central Google ученый
Ван Х. П., Фан Дж. Молекулярная генетика образования сверхкомплектных зубов. Бытие. 2011; 49 (4): 261–77.
CAS PubMed PubMed Central Google ученый
Бей М. Молекулярная генетика развития зубов. Curr Opin Genet Dev. 2009. 19 (5): 504–10.
CAS PubMed PubMed Central Google ученый
Liu X, Li N, Zhang H, Liu J, Zhou N, Ran C, Chen X, Lu Y, Wang X, Qin C, Xiao J. Инактивация Fam20b в мезенхиме нервного гребня мышь вызывает множественные черепно-лицевые дефекты.Eur J Oral Sci. 2018; 126 (5): 433–6.
CAS PubMed PubMed Central Google ученый
Juuri E, Saito K, Ahtiainen L, Seidel K, Tummers M, Hochedlinger K, Klein OD, Thesleff I, Michon F. Стволовые клетки Sox2 + вносят вклад во все эпителиальные клоны зуба через предшественников Sfrp5 +. Dev Cell. 2012. 23 (2): 317–28.
CAS PubMed PubMed Central Google ученый
Juuri E, Jussila M, Seidel K, Holmes S, Wu P, Richman J, Heikinheimo K, Chuong CM, Arnold K, Hochedlinger K, Klein O. Sox2 отмечает способность эпителия образовывать зубы у млекопитающих и рептилий. Разработка. 2013. 140 (7): 1424–32.
CAS PubMed PubMed Central Google ученый
Теслефф И., Ниеминен П. Морфогенез зубов и дифференцировка клеток. Curr Opin Cell Biol. 1996. 8 (6): 844–50.
CAS PubMed Google ученый
Jussila M, Thesleff I. Сигнальные сети, регулирующие органогенез и регенерацию зубов, а также спецификация клонов мезенхимальных и эпителиальных клеток зубов. CSH перспектива биол. 2012; 4 (4): a008425.
Google ученый
Lan Y, Jia S, Jiang R. Молекулярный паттерн зубных рядов млекопитающих. Semin Cell Dev Biol. 2014; 25: 61–70.
PubMed Google ученый
Du W, Du W, Yu H. Роль факторов роста фибробластов в развитии зубов и обновлении резцов. Stem Cell Int. 2018; 2018: 7549160.
Google ученый
Кляйн О.Д., Миновада Г., Петеркова Р., Кангас А., Ю. Б.Д., Лесот Н., Петерка М., Джернвалл Дж., Мартин Г.Р. Гены ростков контролируют развитие диастемных зубов посредством двунаправленного антагонизма эпителиально-мезенхимальной передачи сигналов FGF. Dev Cell. 2006; 11 (2): 181–90.
CAS PubMed PubMed Central Google ученый
Jackman WR, Davies SH, Lyons DB, Stauder CK, Denton-Schneider BR, Jowdry A, Aigler SR, Vogel SA, Stock DW. Манипуляция передачей сигналов Fgf и Bmp у костистых рыб указывает на потенциальные пути эволюционного происхождения многостворчатых зубов. Evol Dev. 2013. 15 (2): 107–18.
CAS PubMed PubMed Central Google ученый
Hokuto I, Perl AK, Whitsett JA. Пренатальное, но не послеродовое ингибирование передачи сигналов рецептора фактора роста фибробластов вызывает эмфизему.J Biol Chem. 2002. 278 (1): 415–21.
PubMed Google ученый
Ли Дж., Ли Л., Тиан Ф., Чжан Л., Сюэ С., Линхардт Р. Дж. Гликозаминогликаномика культивируемых клеток с использованием быстрого и чувствительного подхода LC-MS / MS. ACS Chem Biol. 2015; 10 (5): 1303–10.
CAS PubMed Google ученый
Сюй Д., Эско Дж. Д.. Демистификация взаимодействий гепарансульфат-белок. Анну Рев Биохим.2014; 83: 129–57.
CAS PubMed Google ученый
Zhang F, Zheng L, Cheng S, Peng Y, Fu L, Zhang X, Linhardt RJ. Сравнение взаимодействий различных факторов роста и гликозаминогликанов. Молекулы. 2019; 24 (18): 3360.
CAS PubMed Central Google ученый
Орниц Д.М., Сюй Дж., Колвин Дж. С., МакИвен Д.Г., МакАртур, Калифорния, Кулиер Ф, Гао Дж., Гольдфарб М.Рецепторная специфичность семейства факторов роста фибробластов. J Biol Chem. 1996. 271 (25): 15292–7.
CAS PubMed Google ученый
Орниц Д.М., Ито Н. Путь передачи сигнала фактора роста фибробластов. Wiley Interdiscip Rev Dev Biol. 2015; 4 (3): 215–66.
CAS PubMed PubMed Central Google ученый
Chu EY, Hens J, Andl T, Kairo A, Yamaguchi TP, Brisken C, Glick A, Wysolmerski JJ, Millar SE.Каноническая передача сигналов WNT способствует развитию молочных плакод и важна для инициации морфогенеза молочных желез. Разработка. 2004. 131 (19): 4819–29.
CAS PubMed Google ученый
Munne PM, Tummers M, Järvinen E, Thesleff I, Jernvall J. Работа с индуктивной мезенхимой: Sostdc1 раскрывает роль дентальной мезенхимы в ограничении индукции зубов. Разработка. 2009. 136 (3): 393–402.
CAS PubMed Google ученый
Ahn Y, Sanderson BW, Klein OD, Krumlauf R. Ингибирование передачи сигналов Wnt с помощью Wise (Sostdc1) и отрицательная обратная связь от Shh контролируют количество зубов и их формирование. Разработка. 2010. 137 (19): 3221–31.
CAS PubMed PubMed Central Google ученый
Järvinen E, Birchmeier W, Taketo MM, Jernvall J, Thesleff I. Непрерывное образование зубцов у мышей индуцируется активированной эпителиальной передачей сигналов Wnt / β-катенина. Proc Natl Acad Sci U S A.2006. 103 (49): 18627–32.
PubMed PubMed Central Google ученый
Wang XP, O’Connell DJ, Lund JJ, Saadi I, Kuraguchi M, Turbe-Doan A, Cavallesco R, Kim H, Park PJ, Harada H, Kucherlapati R. Подавление Apc передачи сигналов Wnt регулирует дополнительные формирование зубов в эмбриогенезе и в зрелом возрасте. Разработка. 2009. 136 (11): 1939–49.
CAS PubMed PubMed Central Google ученый
Xavier GM, Patist AL, Healy C, Pagrut A, Carreno G, Sharpe PT, Martinez-Barbera JP, Thavaraj S, Cobourne MT, Andoniadou CL. Активированная передача сигналов WNT в постнатальных SOX2-положительных стволовых клетках зубов может управлять образованием одонтомы. Научный доклад 2015; 5: 14479.
CAS PubMed PubMed Central Google ученый
Попа Е.М., Бухтова М., Такер А.С. Оживление рудиментарной замены зубного ряда у мыши. Разработка. 2019; 146 (3): dev171363.
PubMed Google ученый
Смит М.М., Фрейзер Г.Дж., Мициадис Т.А. Зубная пластинка как источник одонтогенных стволовых клеток: эволюционное происхождение и контроль развития образования зубов в гнатостомах. J Exp Zool Part B. 2009; 312 (4): 260–80.
Google ученый
Чавес М.Г., Ху Дж., Зайдель К., Ли К., Чжон А., Наво А., Хорст О., Кляйн О.Д. Выделение и культура зубных эпителиальных стволовых клеток из резцов взрослой мыши.J Vis Exp. 2014; 87: e51266.
Google ученый
Li J, Feng J, Liu Y, Ho TV, Grimes W, Ho HA, Park S, Wang S, Chai Y. Сигнальная сеть BMP-SHH контролирует судьбу эпителиальных стволовых клеток посредством регуляции своей ниши в развивающийся зуб. Dev Cell. 2015; 33 (2): 125–35.
PubMed PubMed Central Google ученый
Sun Z, Yu W, Navarro MS, Sweat M, Eliason S, Sharp T, Liu H, Seidel K, Zhang L, Moreno M, Lynch T.Sox2 и Lef-1 взаимодействуют с Pitx2, чтобы регулировать развитие резцов и обновление стволовых клеток. Разработка. 2016. 143 (22): 4115–26.
CAS PubMed PubMed Central Google ученый
Харада Х, Мицуясу Т, Тойоно Т, Тойосима К. Эпителиальные стволовые клетки в зубах. Стоматология. 2002; 90 (1): 0001–6.
Google ученый
Coutu DL, Galipeau J. Роль передачи сигналов FGF в самообновлении стволовых клеток, старении и старении.Старение. 2011; 3 (10): 920.
CAS PubMed PubMed Central Google ученый
Volckaert T, Yuan T, Chao CM, Bell H, Sitaula A, Szimmtenings L, El Agha E, Chanda D, Majka S, Bellusci S, Thannickal VJ. Эпителиально-мезенхимальные перекрестные помехи Fgf10-Hippo поддерживают и привлекают базальные стволовые клетки легких. Dev Cell. 2017; 43 (1): 48–59.
CAS PubMed PubMed Central Google ученый
Lombaert IM, Abrams SR, Li L, Eswarakumar VP, Sethi AJ, Witt RL, Hoffman MP. Комбинированная передача сигналов KIT и FGFR2b регулирует экспансию эпителиальных предшественников во время органогенеза. Stem Cell Rep. 2013; 1 (6): 604–19.
CAS Google ученый
Sanz-Navarro M, Seidel K, Sun Z, Bertonnier-Brouty L, Amendt BA, Klein OD, Michon F. Пластичность в нише обеспечивает поддержание популяции Sox2 + стволовых клеток в резце мыши.Разработка. 2018; 145 (1): dev155929.
PubMed PubMed Central Google ученый
Пикфорд К.Э., Холли Р.Дж., Раштон Дж., Ставридис М.П., Уорд К.М., Мерри К.Л. Специфические гликозаминогликаны модулируют нервную спецификацию эмбриональных стволовых клеток мыши. Стволовые клетки. 2011. 29 (4): 629–40.
CAS PubMed Google ученый
Kraushaar DC, Dalton S, Wang L.Гепарансульфат: ключевой регулятор судьбы эмбриональных стволовых клеток. Biol Chem. 2013. 394 (6): 741–51.
CAS PubMed PubMed Central Google ученый
Патель В.Н., Нокс С.М., Ликар К.М., Латроп К.А., Хоссейн Р., Эфтехари С., Уайтлок Дж. М., Элькин М., Влодавский И., Хоффман М.П. Гепараназное расщепление перлекана гепарансульфата модулирует активность FGF10 во время морфогенеза ветвления подчелюстных желез ex vivo. Разработка. 2007. 134 (23): 4177–86.
CAS PubMed Google ученый
Qu X, Carbe C, Tao C, Powers A, Lawrence R, van Kuppevelt TH, Cardoso WV, Grobe K, Esko JD, Zhang X. Развитие слезной железы и передача сигналов Fgf10-Fgfr2b контролируются 2- О- и 6-О-сульфатный гепарансульфат. J Biol Chem. 2011. 286 (16): 14435–44.
CAS PubMed PubMed Central Google ученый
Li Y, Sun C, Yates EA, Jiang C, Wilkinson MC, Fernig DG.Предпочтение связывания гепарина и структуры в семействе факторов роста фибробластов параллельны их эволюционной диверсификации. Откройте Биол. 2016; 6 (3): 150275.
PubMed PubMed Central Google ученый
Lindahl U, Li JP. Взаимодействие между гепарансульфатом и белками — дизайн и функциональное значение. Int Rev Cel Mol Biol. 2009. 276: 105–59.
Google ученый
Li JP, Kusche-Gullberg M. Гепарансульфат: биосинтез, структура и функция. Int Rev Cell Mol Biol. 2016; 325: 215–73.
CAS PubMed Google ученый
Льюис А.Е., Васудеван Х.Н., О’Нил А.К., Сориано П., Буш Дж. Широко используемый трансген Wnt1-Cre вызывает фенотипы развития за счет эктопической активации передачи сигналов Wnt. Dev Biol. 2013. 379 (2): 229–34.
CAS PubMed PubMed Central Google ученый
Nguyen H, Rendl M, Fuchs E. Tcf3 управляет функциями стволовых клеток и подавляет определение судьбы клеток в коже. Клетка. 2006; 127 (1): 171–83.
CAS PubMed Google ученый
Perl AK, Wert SE, Nagy A, Lobe CG, Whitsett JA. Раннее ограничение периферических и проксимальных клонов клеток во время формирования легкого. Proc Natl Acad Sci. 2002. 99 (16): 10482–7.
CAS PubMed Google ученый
Мадисен Л., Цвингман Т.А., Санкин С.М., Оу С.В., Заривала Х.А., Гу Х., Нг Л.Л., Палмитер Р.Д., Гаврилич М.Дж., Джонс А.Р., Лейн ES. Надежная и высокопроизводительная система Cre отчетов и характеристик для всего мозга мыши. Nat Neurosci. 2010. 13 (1): 133–40.
CAS PubMed Google ученый
Maretto S, Cordenonsi M, Dupont S, Braghetta P, Broccoli V, Hassan AB, Volpin D, Bressan GM, Piccolo S. Картирование передачи сигналов Wnt / β-катенина во время развития мышей и в колоректальных опухолях.Proc Natl Acad Sci. 2003. 100 (6): 3299–304.
CAS PubMed Google ученый
Бай С.Б., Ауэрбах В., Ли Дж.С., Стивен Д., Джойнер А.Л. Gli2, но не Gli1, необходим для начальной передачи сигналов Shh и эктопической активации пути Shh. Разработка. 2002. 129 (20): 4753–61.
CAS PubMed Google ученый
Гудрич Л.В., Миленкович Л., Хиггинс К.М., Скотт М.П.Измененные судьбы нервных клеток и медуллобластома у мутантов с заплатой у мышей. Наука. 1997. 277 (5329): 1109–13.
CAS PubMed Google ученый
Arnold K, Sarkar A, Yram MA, Polo JM, Bronson R, Sengupta S, Seandel M, Geijsen N, Hochedlinger K. Взрослые стволовые клетки и клетки-предшественники Sox2 + важны для регенерации тканей и выживания мышей. Стволовая клетка клетки. 2011; 9 (4): 317–29.
CAS PubMed PubMed Central Google ученый
Shaham O, Smith AN, Robinson ML, Taketo MM, Lang RA, Ashery-Padan R. Pax6 необходим для дифференцировки клеток волокна хрусталика. Разработка. 2009. 136 (15): 2567–78.
CAS PubMed PubMed Central Google ученый
Jing D, Zhang S, Luo W, Gao X, Men Y, Ma C, Liu X, Yi Y, Bugde A, Zhou BO, Zhao Z. Очистка твердых и мягких тканей органов с помощью Метод PEGASOS. Cell Res. 2018; 28 (8): 803–18.
CAS PubMed PubMed Central Google ученый
Wu J, Feng JQ, Wang X. Гибридизация in situ на парафиновых срезах мышей с использованием DIG-меченых РНК-зондов. В: Odontogenesis 2019. Нью-Йорк: Humana Press; 2019. стр. с163–71.
Google ученый
Wu J, Wang X. Полная гибридизация in situ мышиных эмбрионов с использованием DIG-меченых РНК-зондов. В: Odontogenesis 2019. Нью-Йорк: Humana Press; 2019. стр. с151–9.
Google ученый
Sun X, Li L, Overdier KH, Ammons LA, Douglas IS, Burlew CC, Zhang F, Schmidt EP, Chi L, Linhardt RJ. Анализ общих дисахаридов гликозаминогликанов в моче человека методом жидкостной хроматографии и тандемной масс-спектрометрии. Anal Chem. 2015; 87 (12): 6220–7.
CAS PubMed PubMed Central Google ученый
Santos-Ocampo S, Colvin JS, Chellaiah A, Ornitz DM. Экспрессия и биологическая активность фактора роста фибробластов мыши-9.J Biol Chem. 1996. 271 (3): 1726–31.
CAS PubMed Google ученый
Zhang Z, McCallum SA, Xie J, Nieto L, Corzana F, Jiménez-Barbero J, Chen M, Liu J, Linhardt RJ. Структуры растворов химически синтезированного гепарина и его предшественников. J Am Chem Soc. 2008. 130 (39): 12998–3007.
CAS PubMed PubMed Central Google ученый
Что такое номера и названия зубов — система нумерации зубов?
Вы запутались цифрами зубов? Вы знаете, что у нас есть нумерация для каждого зуба?
Я — «если я положу мертвый зуб обратно под подушку, Бог вернет его обратно?» Тогда почему я не получаю его обратно.”
Мама: «Не волнуйтесь, вы получите больше, чем имеете. Имейте немного веры. Зуб феи подарит тебе лишние зубы ».
Мы росли, слушая сказку о зубах.
Все зубы выглядят по-разному.
Каждый зуб встроен в кость, называемую лункой, с внутренней частью, называемой корнем. Видимая часть зуба называется коронкой.
Зубы во рту расположены особым образом.
Сколько зубов у человека?
Во рту есть четыре класса зубов.
Кусаем по-разному для разных блюд.
Для того, чтобы съесть гамбургер, мы используем восемь передних зубов с острыми краями, как ножницы, таким образом известные как резцы. Они помогают кусать и резать пищу. В каждой челюсти по четыре резца. Всего на обеих челюстях восемь резцов.
Во время еды в KFC мы используем острые зубы, расположенные рядом с резцами, известные как клыки. В каждой челюсти по два клыка. Итого у нас четыре клыка. Помогает разрывать пищу, особенно мясо.
Мы кладем овес или любой другой мягкий пищевой продукт прямо на одну сторону рта и начинаем его растирать и измельчать. Этот способ измельчения, затирания, кусания облегчается премолярами и коренными зубами. Премоляры немного меньше моляров, в каждой челюсти по четыре премоляра. Всего у взрослых бывает восемь премоляров. У детей их нет. Они известны потому, что предшествуют коренным зубам.
Коренные зубы — это зубы на самом дальнем конце. Моляры имеют максимальную площадь поверхности и широкие.большой бугорок на молярах не только помогает при измельчении пищи, но и при раздавливании. В каждой челюсти по шесть коренных зубов. Всего на обеих челюстях по 12 коренных зубов.
Первый набор зубов у ребенка известен как молочные зубы или молочные зубы. К раннему подростковому возрасту 11-12 лет эти молочные зубы отпали и были заменены постоянными. По мере того как человек стареет, эти постоянные зубы также выпадают, но никакие другие зубы не заменяют их.
Молочные зубы известны как молочные зубы, в то время как зубы у взрослых постоянные, поэтому они называются постоянными зубными рядами.
Верхние зубы расположены дугообразно, как и нижние.
Верхние и нижние зубы разделены на четыре части.
Стоматологическая формула
Стоматологическая формула — это формула, разработанная для легкого запоминания количества и типа зубов у детей и взрослых.
Зубная формула включает только одну сторону каждой челюсти и аналогична другой.
Каждая челюсть разделена на две части по средней линии.Он представляет собой только одну сторону верхней или нижней челюсти.
Число и названия зубов
Резцов — 4 шт. У них плоская поверхность и острый горизонтальный край, позволяющий кусать и разрезать пищу.
Есть два клыка. По одному с обеих сторон от резцов для разрыва пищи. Самые активные зубы с самыми длинными корнями.
Премоляров. 4 премоляра позади клыка, помогающие жевать.
Есть шесть коренных зубов. Они используются для измельчения пищи перед проглатыванием.
У каждого зуба есть номинал, который представлен первой буквой его названия.
Например, резцы по I, клыки по C, премоляры по P, моляры по M.
Если рассматривать половину челюсти, то получается зубная формула
Два резца
Одна собака
Два премоляра
Три моляра
Стоматологическая формула становится 2123
Таких челюстей четыре. Общее количество зубцов становится 32.
У детей младше 12 лет нет премоляров 2102, поэтому общее количество зубов у ребенка составляет 20
Горизонтальная линия — это линия, символизирующая рот. Вверху по строке числитель — это зубы верхней или верхней челюсти, а под линией знаменателя — нижние или нижнечелюстные зубы.
В возрасте 19-22 лет формируется последний коренной зуб, который сопровождается сильной болью в задней части челюсти. Интересным фактом является то, что он известен как зуб мудрости.Не потому, что они мудрее, а потому, что развиваются последними.
Это годы, когда человек достигает высшей мудрости.
Человек за всю жизнь вырастает два набора зубов.
Система нумерации зубьев
Большинство стоматологов путаются с системой нумерации зубов.
Существует зубная формула для зубов.
Мне делали стоматологическое лечение, и по какой-то причине мне пришлось ехать. Теперь, чтобы завершить лечение, стоматолог должен знать, над какими зубами ему нужно работать.
Очень важно видеть номера зубов, чтобы вести надлежащую запись.
У нас есть три системы нумерации зубов, пользующиеся наибольшим всеобщим доверием
Горизонтальная линия разделяет верхнюю и нижнюю челюсти.
Вертикальная линия делится на правую и левую от средней линии
. Ваше право — пациенты слева, и наоборот.
Чтобы устранить неисправность, поставьте знак «плюс», во втором квадранте образуется буква «L». Итак, второй квадрант — это оставшиеся пациенты.В любой из систем нумерации зубов правая сторона верхней челюсти запускается первой.
Универсальная нотариальная система
Универсальная система нумерации временных зубов
Алфавиты (A-Z) используются в верхнем регистре. Рисуются как горизонтальные, так и вертикальные линии, начиная с
. Универсальная система нумерации временных зубовмакс. Правый квадрант от последнего зуба, равного
2-й молочный моляр как A
1-й молочный моляр как B
собак как C
боковой резец по D
центральный резец по E
продолжайте буквы до левого квадранта верхней челюсти от центра до моляра
Номера зубов в квадранте левой верхней челюсти
центральный резец как F
сбоку как G
собака как H
1-й моляр как I
2-й моляр как J
Вместо того, чтобы начинать с правого квадранта.начинаем нижнюю челюсть от левого квадранта от моляров до центра
Левый квадрант нижней челюсти
2-й молочный корень как K
1-й молочный моляр как L
клык как M
боковой резец по N
центральный резец по O
для правого квадранта нижней челюсти от центра до моляров
центральный резец как P
боковой как Q
собак как R
1-й моляр как S
2-й моляр как T
Так в универсальной системе для лиственных пород мы отмечаем от Ato T.начиная с правого квадранта верхней челюсти, переходя к левому квадранту верхней челюсти, затем переходя к левому квадранту нижней челюсти и, наконец, к правой нижней челюсти.
Универсальная система нумерации постоянных зубов
Универсальная система нумерации для временных зубовВ молочном алфавите используются, а в постоянном прикусе используются числа.
Номера зубов от правый квадрант верхней челюсти
3-й моляр как «1»
2-й моляр как «2»
1-й моляр как «3»
2-й премоляр как «4»
1-й премоляр как «5»
клык как «6»
боковой резец по «7»
центральный резец по восьмерке
продолжить нумерацию зубьев до левый размер
центральный резец по «9»
боковой резец на ’10’
собак как ’11’
1-й премоляр ’12’
2-й премоляр как ’13’
1-й моляр как ’14’
2-й моляр как ’15’
3-й моляр как ’16’
начинаем с номера зубов нижней челюсти с левой стороны
3-й моляр как ’17’
2-й моляр как ’18’
1-й моляр как ’19’
2-й премоляр как ’20’
1-й премоляр как ’21’
клык как ’22’
боковой резец на ’23’
центральный резец на ’24’
переходим к правый квадрант нижней челюсти
центральный резец на ’25’
боковой резец по ”
собак как ’27’
1-й премоляр как ’28 ‘
2-й премоляр как ’29’
1-й моляр как ’30’
2-й моляр как ’31 ′
3-й моляр как ’32’
Система Зигмонди и Пламер
назван в честь конструктора
Алфавиты фиксируются для типа зуба.
так все квадранты для лиственных
центральный молочный резец всех квадрантов по ‘A’
латеральный молочный резец всех квадрантов по’B ’
лиственных клыков всех квадрантов как «C»
1-й молочный моляр во всех квадрантах как «D»
2-й молочный моляр всех квадрантов как ’E’
Соотнесите пальмер «P» со знаком плюс.
В постоянном прикусе номера зубов используются вместо букв и фиксируются для всего квадранта
центральный резец всех квадрантов ‘1’
боковых резцов всех квадрантов на «2»
клыков всех квадрантов как «3»
1-й премоляр всех квадрантов как «4»
2-й премоляр всех квадрантов как «5»
1-й моляр всех квадрантов как «6»
2-й моляр всех квадрантов как «7»
3-й моляр всех квадрантов как ’8 ′
, например, левый клык верхней челюсти обозначен цифрой «3».
Система прямых иностранных инвестиций
Федерация Dentaire International принята ВОЗ. Это также известно как двузначная система.
Знак плюс не используется. Так что это хорошо, когда вы хотите хранить данные на компьютере.
Зубу присвоены две цифры. Первая цифра представляет квадрант, а вторая цифра представляет зуб.
Постоянный прикус 1, 2, 3, 4.
правый квадрант верхней челюсти как «1»
левый квадрант верхней челюсти как ’2 ′
Левый квадрант нижней челюсти как ’3 ′
Правый квадрант нижней челюсти как ‘4’
аналогично для молочных зубов
правый квадрант верхней челюсти как «5»
левый квадрант верхней челюсти как «6»
Левый квадрант нижней челюсти как ’7 ′
нижнечелюстной правый квадрант как ’8 ′
Правая центральная челюсть 51 «пять к одному», а не пятьдесят один.Точно так же левый клык верхней челюсти имеет размер 73 «семь-три», а не семьдесят три.
Для правого квадранта верхней челюсти для временных зубов.
все зубья начинаются с ’5 ′
центральный резец как ’51 ′
сбоку как ’52’
собак как ’53’
1-й моляр как ’54’
2-й моляр как ’55’
Нумерация зубов левого квадранта верхней челюсти
центральный резец как ’61’
сбоку как ’62’
собак как ’63’
1-й моляр как ’64’
2-й моляр как ’65’
7 — это номер левого квадранта нижней челюсти.
центральный резец как ’71’
сбоку как ’72’
собак как ’73’
1-й моляр как ’74 ′
2-й моляр как ’75’
Номера зубов для правого квадранта нижней челюсти
центральный резец как ’81’
сбоку как ’82’
собак как ’83’
1-й моляр как ’84’
2-й моляр как ’85’
Система нумерации зубов FDI для постоянного прикуса
квадранта делятся на 1,2,3,4
Число зубов правого квадранта верхней челюсти
центральный резец as’11 ’
сбоку как ’12’
собак как ’13’
1-й премоляр как ’14’
2-й премоляр как ’15’
1-й моляр как ’16’
2-й моляр как ’17’
3-й моляр как ’18’
Номера зубов левого квадранта верхней челюсти
центральный резец как ’21’
сбоку как ’22’
собак как ’23’
1-й премоляр как ’24’
2-й премоляр как ’25’
1-й моляр как ’26’
2-й моляр как ’27’
3-й моляр как ’28’
Нумерация зубов для левого квадранта нижней челюсти
центральный резец как ’31’
сбоку как ’32’
собак как ’33’
1-й премоляр как ’34’
2-й премоляр как ’35’
1-й моляр как ’36’
2-й моляр как ’37’
3-й моляр как ’38’
для правого квадранта нижней челюсти
центральный резец как ’41’
сбоку как ’42’
собак как ’43’
1-й премоляр как ’44’
2-й премоляр как ’45’
1-й моляр как ’46’
2-й моляр как ’47’
3-й моляр как ’48’
Я рекомендую вам попрактиковаться в системе нумерации зубов.